Половой диморфизм функциональной организации мозга при обработке речевой информации
Н.В. Вольф, О.М. Разумникова
Государственное учреждение НИИ физиологии
Сибирского отделения РАМН
Методические достижения последних десятилетий, позволившие проводить прижизненные исследования, как структуры, так и функций полушарий мозга у здоровых испытуемых, убедительно показали, что половые различия в полушарной организации когнитивных функций не только связаны с особенностями обусловленной полом социализации, но имеют под собой биологическую основу. Половые различия проявляются не только и, возможно, не столько в успешности осуществления некоторых видов мыслительной деятельности, но и в стратегиях, на которых она базируется. Нейрофизиологических исследования последних лет все больше направлены на специальное изучение механизмов мозга, определяющих связанную с полом специфику когнитивных процессов. Однако накопившиеся факты рассматриваются в основном в рамках гипотезы, которая связывает половые различия в когнитивных процессах с разной выраженностью функциональной асимметрии мозга у особей мужского и женского пола. Эта точка зрения была аргументирована в обзоре МакГлоун (J. McGloun, 1980) на основании довольно полного анализа имевшихся к тому времени экспериментальных данных. Вместе с тем современные знания о полушарной организации вербальных процессов свидетельствуют, что в отличие от фонетического анализа, связанного преимущественно с левым полушарием мозга, в анализе семантических и лингвистических характеристик речевого материала значительная роль принадлежит правому полушарию. В связи с этим на первое место выходит вопрос не о разделении функций между полушариями, а о том, каким образом каждое из полушарий принимает участие в осуществлении конкретной речевой деятельности и как при этом осуществляются процессы межполушарного взаимодействия.
Именно с этих позиций мы попытаемся рассмотреть современные данные о половом диморфизме в полушарной организации речевых функций и возможности их интерпретации.
Половые различия обнаружены в величине planum temporale, области вовлеченной в высшие речевые функции и входящей в состав зоны Вернике. По данным последних исследований с использованием метода ядерного-магнитного резонанса (ЯМР) выраженность анатомической асимметрии, характеризующейся большей величиной planum temporale в левом полушарии у мужчин по сравнению с женщинами (J.J. Kulynych et al., 1994; D.R. Weinberger, 1994; C.D Good et al., 2001; J. Shapleske et al., 1999). В противоположность этому посмертное исследование срезов мозга показало большую величину областей, ассоциируемых с областями Вернике и Брока, в левом полушарии у женщин по сравнению с мужчинами (J. Harasty et al., 1997), что, однако, противоречит данным более раннего аналогичного исследования (Wada, 1975). Недавно установлено, что половые различия касаются не только величины, но и клеточной организации левой planum temporale. В ряде исследований большая плотность серого вещества в речевых зонах обнаружена у женщин по сравнению с мужчинами (E.T. Schlaepfer et al., 1995; S.F. Witelson et al., 1995; R.C. Gur et al., 1999). Часто рассматриваемой характеристикой при изучении асимметрии речевых зон является морфология Сильвиевой борозды в левом и правом полушариях. Для ее горизонтальной части и Сильвиевой борозды в целом обнаружена большая асимметрия у праворуких мужчин по сравнению с женщинами и леворукими мужчинами (S.F. Witelson, D.L. Kigar, 1992). Эти различия остались достоверными и с учетом поправок, учитывающих разницу в весе мозга. Однако, в другой работе, также проведенной с использованием метода ЯМР, не выявлено половых различий в асимметрии сильвиевой борозды (A.L. Foundas et al., 1999). С применением метода ЯМР было установлено, что, у праворуких мужчин и леворуких женщин париетальная область левого полушария больше, чем правого. В отличие от этого у леворуких мужчин и праворуких женщин асимметрия отсутствовала (A. Kertesz et al., 1990).
Во многих исследованиях обнаружены половые различия в морфометрических показателях мозолистого тела – структуры, связывающей два полушария мозга. У женщин выявлена большая ширина каудальной части мозолистого тела в области перешейка и валика и больший относительный размер этой части мозолистого тела (W. Byne, et al., 1988; J.M. Clarke et al., 1989; S.F. Witelson, 1989; R.L. Holloway et al., 1993; H. Steinmetz et al., 1992; J.M. Clarke, E. Zaidel, 1994), а также больший общий размер мозолистого тела (R.L. Holloway, M.C. de Lacoste, 1986). Эти данные наиболее бесспорны в отношении перешейка, тогда как не все исследования подтверждают наличие половых различий в величине сплениума мозолистого тела (W. Byne, R. Bleier, L. Houston, 1988; S. Clarke et al., 1989; A. Prokop, M. Oehmichen, K. Zilles, 1990). Некоторые авторы считают, что для сплениума более стабильные результаты дает изучение формы. Так, исследования Кларка с соавторами (S. Clarke et al., 1989) и проведенный ими ретроспективный анализ данных других лабораторий свидетельствуют, что эта часть мозолистого тела у женщин является более выпуклой, чем у мужчин. Что касается величины сплениума, то большинство данных подтверждает существование тенденции к увеличению его размера у женщин по сравнению с мужчинами, однако половые различия стабильно выявляются только в исследованиях с учетом таких параметров, как величина мозолистого тела и мозга в целом, возраст, право- и леворукость испытуемых (A.B. Wisniewski, 1998). В то же время даже при невыявленных различиях в величине речевых зон и размерах мозолистого тела показано, что у мужчин, но не у женщин, полушарные различия в размерах задних речевых зон (сильвиевой борозды и planum temporale) отрицательно коррелируют с площадью поперечного сечения мозолистого тела в области isthmus: более выраженная анатомическая асимметрия полушарий сочетается с уменьшением числа каллозальных связей (F. Aboitiz et al., 1992; E.Zaidel et al., 1995). Высказывается точка зрения, что менее латерализованный мозг нуждается в большем количестве межполушарных связей (S.F. Witelson, 1985; 1989; M. Habib et al., 1991). Однако не исключена вероятность того, что большее развитие мозолистого тела при симметрии полушарий обусловлено особенностями их онтогенетического развития (одновременное созревание) (S. Clarke, E. Zaidel, 1994).
Половые различия, обнаруженные в морфометрии задней части мозолистого тела, которая содержит волокна от каудальных речевых областей, могут быть дополнительным фактором, обусловливающим различия в полушарной организации речевых функций у мужчин и женщин (S.F. Witelson, 1989). Это подтверждается данными о взаимодействии факторов пола и "рукости" в морфометрических исследованиях. У мужчин-правшей область перешейка меньше, чем у мужчин с невыраженным преимуществом одной из рук, у женщин не наблюдается связи величины этой области мозолистого тела с "рукостью" (S.F. Witelson,1989; V.H. Denenberg et al., 1991; M. Habib et al., 1991; S. Clarke, E. Zaidel, 1994). Исследование корреляций между показателями асимметрии в задании, требующем лексического решения, и размерами мозолистого тела также показало, что только у мужчин величина перешейка отрицательно коррелирует с полушарными различиями в речевых функциях, связанных с каудальными речевыми зонами. Заслуживает внимания и тот факт, что у мужчин в отличие от женщин обнаружена также отрицательная корреляция между эффективностью воспроизведения слогов, предъявлявшихся в правое ухо при дихотическом тестировании, и размером передней части мозолистого тела, для которой не выявлено морфометрических половых различий (S. Clarke, E. Zaidel, 1994). В исследованиях на испытуемых разного пола установлено, что только у мужчин преимущество правого поля зрения в обработке ассоциированных слов отрицательно коррелировало с площадью сечения перешейка мозолистого тела (E. Zaidel et al., 1995). У женщин показатель беглости речи (вычисленный на основе показателей способности продуцировать слова, начинающиеся на заданную букву, подбирать слова одинакового значения и составлять предложения, со словами, с определенными начальными буквами) был позитивно связан с величиной задней части мозолистого тела, определенной методом ЯМР (M. Hines et al., 1992). Таким образом, разные вербальные процессы обнаруживают корреляцию с размерами разных отделов мозолистого тела, и эти зависимости отличаются у мужчин и женщин.
Новые данные о половых различиях в представительстве речевых функций внутри левого полушария были получены в исследованиях Кимуры (D. Kimura 1980, 1983). После поражений передних корковых областей левого полушария, преимущественно захватывавших зону Брока, наблюдались сходные афазические расстройства у мужчин и женщин. Однако при более каудальных поражениях, в основном связанных с зоной Вернике, выявлены существенные половые различия: у мужчин случаи афазии встречались значительно чаще, чем у женщин. На основе этих наблюдений был сделан вывод о различиях внутриполушарной организации речевых функций у мужчин и женщин: у женщин продуктивные речевые функции соотносятся с передними областями левого полушария, тогда как у мужчин в речевых процессах участвуют, как передние, так и задние отделы левого полушария, причем роль последних весьма существенна. Этот вывод впоследствии нашел подтверждение в клинических исследованиях влияния на речевые функции стимуляции передних и задних корковых областей у мужчин и женщин (C. Mateer et al., 1982; G.A. Ojemann 1983; G.A. Ojemann et al., 1989). Аналогичные данные были получены и на здоровых испытуемых с применением метода мануально-вербальной интерференции (R.S. Lewis, L. Christiansen, 1989).
Таким образом, результаты большинства исследований свидетельствуют о том, что латеральные особенности морфологии речевых зон более выражены у мужчин, чем у женщин. Этот факт часто используют как одно из подтверждений большей эквипотенциальности полушарий у женщин по сравнению с мужчинами. Однако данные о половых различиях внутриполушарной организации речевых функций, морфофункциональной организации мозолистого тела, взаимодействии факторов пола и "рукости" в морфометрических исследованиях свидетельствуют против такой упрощенной трактовки. В частности, половые различия в представленности речевых функций вдоль оси "лоб-затылок" в левом полушарии могут определять особенности межполушарного взаимодействия, степень и характер вовлечения каждого из полушарий и их отделов в осуществление конкретных речевых функций. Подтверждением этого служат приведенные выше данные сопоставления результатов психофизиологического тестирования при разных речевых заданиях с данными морфологии передних и задних отделов мозолистого тела у мужчин и женщин (M. Hines et al., 1992; S. Clarke, E. Zaidel, 1994). При этом уменьшение латеральных морфологических различий полушарий у женщин может быть объяснено не только их эквипотенциальностью, но изменением соотношения левополушарных и правополушарных стратегий.
Следовательно, при изучении половых различий в полушарной организации речевых функций необходимо учитывать характер осуществляемой вербальной деятельности. Представление о полушарной организации этой деятельности может быть получено только на основе комплексного анализа как межполушарных, так и внутриполушарных функциональных взаимодействий.
В современных психофизиологических исследованиях восприятие речи рассматривают как многоступенчатый процесс, в котором уровень фонетического анализа является первым надсенсорным символическим уровнем речевого анализа (Л.Я. Балонов с соавт., 1985). Основная масса исследований функциональной асимметрии мозга относится именно к этому уровню, данных о полушарной организации других уровней языка значительно меньше.
В условиях угнетения одного полушария после унилатеральных электросудорожных припадков было показано, что повышение порогов обнаружения звуков речи, ухудшение опознания фонем, слогов и слов наблюдается только при инактивации левого полушария и обусловлено утратой смыслоразличительной функции фонем. Структурами правого полушария не производится фонематический анализ, значение слова извлекается из целостного звукового образа (Л.Я. Баллонов, В.Л. Деглин, 1976; Л.Я. Баллонов и др., 1985). Эти данные в целом совпадают с результатами исследований, проведенных на здоровых людях с использованием метода дихотического тестирования, в которых показано преимущество левого полушария в восприятии бессмысленных слогов и слов (С. Спрингер, Г. Дейч, 1983; И.А. Вартанян с соавт., 1988; M.P. Bryden, 1988).
Лишь немногочисленные данные позволяют сравнивать полушарную организацию фонетического анализа речи у мужчин и женщин. Так, превалирование женщин среди больных, проходивших курс электросудорожной терапии, (119 из 150 больных, Л.Я. Балонов, В.Л. Деглин, 1976, стр. 40) позволяет заключить, что фонетический анализ у женщин, связан с левым полушарием мозга. Исследование влияния асинхронности предъявления слогов в дихотической паре на эффективность их воспроизведения у детей различных возрастных групп выявило половые различия, обусловленные лучшим восприятием опережающих слогов девочками при увеличении асинхронности, что может отражать большую скорость фонетического анализа (P. Mirabile et al., 1977). При анализе результатов дихотических исследований латерализации восприятия слогов, состоящих из согласной и гласной, и использовавших в качестве сравниваемых показателей средние значения разницы воспроизведения с правого и левого уха, только в одной работе выявлен больший эффект правого уха у женщин по сравнению с мужчинами (Dorman, R.J. Porter, 1975 цит. по J. McGlone, 1980), в большинстве исследований, не обнаружено половых различий (J. McGlone, W. Davidson, 1973; M. Hiscock, M. MacKay, 1985; M.P. Bryden, 1988; S. Clarke, E. Zaidel, 1994). В отличие от этого Д. А. Лейк и М.Р.Брайден (D.A.Lake, M.P. Bryden, 1976) обнаружили, что у мужчин преимущество правого уха выражено сильнее, чем у женщин. Сравнение мужчин и женщин по такому показателю, как количество испытуемых, показавших эффект правого уха, выявило как достоверно большую встречаемость левостороннего доминирования у мужчин (M.P. Bryden,1973; D.A. Lake, M.P. Bryden, 1976), так и отсутствие половых различий (M.P. Bryden, 1975;M.P. Bryden et al., 1983). Тем не менее, обзор исследований половых различий латерализации при выполнении дихотического теста, сделанный в двух работах (M. Hiscock, M. MacKay, 1985; M.P. Bryden, 1988), привел авторов к выводу, что половые различия невелики и выявляют лишь тенденцию большего преимущества правого уха у мужчин по сравнению с женщинами при предъявлении бессмысленных речевых сигналов.
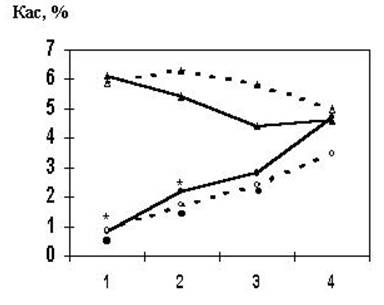
В последнее время появились данные, которые указывают по крайней мере на одну из возможных причин вариативности результатов оценки половых различий в фонетическом анализе дихотически предъявляемых слогов. Обнаружено, что на начальном этапе прослушивания длинного (120 пар) списка слогов у женщин наблюдается лучшее воспроизведение информации с левого, а у мужчин - с правого уха. По мере выполнения задания показатели асимметрии обеих групп сближались (B.E. Wexler, E. Lipman, 1988; P. Munro, E. Govier, 1993). Предполагается, что причиной такого эффекта могут быть половые различия в реакции на новизну информации: использование мужчинами преимущественно реакции приближения к стимулу (левополушарной стратегии), а женщинами – стратегии избегания (правополушарной) (B.E. Wexler, E. Lipman, 1988). Чтобы проверить это предположение мы исследовали временную динамику воспроизведения дихотически предъявляемых слогов (120 пар) у мужчин (65 человек) и женщин (52 человека) в двух экспериментальных сериях. После предъявления каждой пары требовалось письменно воспроизвести оба слога. Оказалось, что и при повторном тестировании у как мужчин, так и у женщин наблюдается та же временная динамика изменения коэффициента асимметрии, как и при первом тестировании (Рис.1). Изменения коэффициента асимметрии были обусловлены нарастанием воспроизведения слогов, адресованных левому полушарию, у женщин и правому - у мужчин. Следовательно, наблюдаемые половые различия не связаны с новизной ситуации тестирования. В то же время проведенные нами эксперименты с избирательным воспроизведением слогов с одного из каналов при дихотическом предъявлении в отличие от ситуации распределенного внимания не выявило половых различий на начальных этапах тестирования. В процессе тестирования у всех испытуемых наблюдалось возрастание воспроизведения слогов с правого уха, более выраженное у женщин (О.М. Разумникова, Н.В. Вольф, 1997). Приведенные данные позволяют связать изменения полушарного вклада в обработку информации в ходе длительного тестирования с половыми особенностями организации процессов внимания.
Рис.1. Зависимость коэффициента асимметрии воспроизведения слогов от длительности тестирования в двух экспериментальных сериях у мужчин и женщин.
Таким образом, различия в длине используемых списков, а также в характере процедуры тестирования могут влиять на проявление половых различий в результатах фонетического анализа при латерализованном предъявлении вербальных стимулов.
Полушарные особенности обработки семантически значимой вербальной информации
Наиболее изучен вопрос о роли полушарий в слуховом восприятии существительных. Их восприятие оценивалось с точки зрения положения на шкале абстрактности - конкретности, образности. Хотя результаты, полученные в этих исследованиях неоднородны, все же преобладающее количество работ свидетельствует об участии в обработке абстрактных существительных преимущественно левого полушария, тогда как при обработке конкретных существительных преимущество левого полушария исчезает (H.D. Ellis, J.W. Sheperd, 1974; J. Day, 1979; Д.А. Кауфман, О.П. Траченко, 1985; P.W. Ely et al., 1989; Т.В. Черниговская с соавт., 1996). Однако влияние образности-конкретности слов на выраженность преимущества правого уха зарегистрировано не во всех исследованиях (H.B. Orenstein, W.B. Meighan, 1976). При дихотическом исследовании с применением в качестве стимульного материала серий по три конкретных слова степень асимметрии у мужчин была выше, чем у женщин. Такой же характер половых различий наблюдался у детей различных возрастных групп (И.А. Вартанян с соавт., 1988). Наряду с эффектом «правого уха», разнообразные комбинации дихотического теста и моторных реакций выявили, что для мужчин в большей мере характерна стабильная асимметрия взаимодействия полушарий, тогда женщины используют разные стратегии переработки речевых сигналов (T. Welsh, D. Elliot, 2001). Однако использование в качестве дихотически предъявляемых стимулов названий чисел, обнаружив эффект правого уха у всех испытуемых, не выявило половых различий в его величине (J. McGlone, W. Davidson, 1973; M.P Bryden, 1975). Таким образом, положение на шкале образности-абстрактности по-разному влияет на проявление функциональной асимметрии в организации вербальных процессов у мужчин и женщин. Очевидно, что уменьшение коэффициента асимметрии может происходить как за счет снижения функциональных возможностей левого, так и за счет повышения функциональных возможностей правого полушария. К сожалению, отсутствие в приведенных работах данных о продуктивности деятельности, связанной с каждым полушарием, не позволяет ответить на этот вопрос.
Существует ограниченное количество данных об особенностях нарушений речевой памяти у мужчин и женщин после локальных повреждений мозга. Так, например, описаны более выраженные нарушения памяти после разрушений в области левой височной доли у мужчин по сравнению с женщинами (M.R. Smith, B. Milner, 1984; M.R. Trenerry et al., 1995). При менее точно локализованных поражениях левого полушария на небольшом контингенте испытуемых описаны как аналогичные результаты (J. McGlone, 1978), так и отсутствие половых различий (W.G. Snow, S.Cheese, 1985).
При анализе продуктивности вербальной памяти в ситуации свободного припоминания словесной информации воспроизведение у женщин было более успешным, чем у мужчин (Р.С. Трубникова, 1974; J. Levy, 1980; Rugg et al., 1989; D.J. Нerrmann et al., 1992; A. Herlitz, 1997.). Эффективность отсроченного воспроизведения слов по ряду данных также выше у женщин по сравнению с мужчинами (Р.С. Трубникова, 1974; С.M. Sinton, 1981; E.B. Jones et al., 1987). Большинство же данных о половых различиях в вербальной памяти было получено как побочный продукт главным образом в исследованиях возрастных особенностей памяти (M.L. Bleecker et al.,1988; D.F. Hulstch et al., 1991; G.J. Larrabee, T.H. Crook, 1993; K.W. Schaie, S.L. Willis, 1993; Zelinski et al., 1993; R.D. Hill et al., 1995; R. Portin et al., 1995; P. Rabbitt et al., 1995) или при разработке нормативных данных для тестов памяти (R. Bolla-Wilson, M.L. Bleeker, 1986; G. Geffer et al., 1990; C.M. Temple, K.M. Cornish, 1993). Во всех этих исследованиях у женщин обнаружены более высокие показатели вербальной памяти, чем у мужчин. Однако физиологическая основа выявленного преимущества, его связь с особенностями полушарной интеграции экспериментально не изучены.
Воспроизведение запоминаемых слов осуществлялось в трех вариантах. При непосредственном воспроизведении испытуемые записывали в произвольном порядке запомненные слова сразу после прослушивания каждого из 9 списков. При отсроченном воспроизведении
после записи слов последнего списка испытуемым неожиданно было предложено воспроизвести все запомненные слова из всех предъявленных списков или дифференцировать нажатием на соответствующие клавиши клавиатуры компьютера среди последовательно предъявляемых на экране монитора слов те, которые имелись или отсутствовали в этих списках (отсроченное узнавание) (Н.В. Вольф, 2000).
При непосредственном воспроизведении слов только у мужчин наблюдалось преимущество левого полушария. При этом различия в эффективности воспроизведения с правого уха (левого полушария) у мужчин и женщин отсутствовали, но женщины воспроизводили больше, чем мужчины, слов с левого уха. Таким образом, уменьшение латеральных различий у женщин обусловлено возрастанием эффективности обработки информации, адресованной правому полушарию (Рис.2). При отсроченном воспроизведении женщины воспроизводили больше слов, чем мужчины как с правого, так и с левого уха. У испытуемых обоего пола отсутствовали латеральные различия в эффективности воспроизведения (Рис.2). При узнавании слов женщины воспроизводили больше, чем мужчины слов с левого уха.
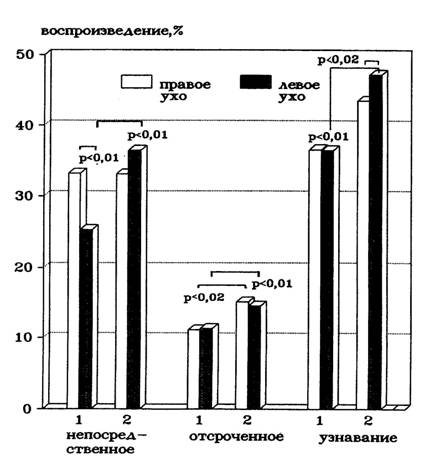
Рис.2. Различия в эффективности воспроизведения дихотически предъявленных списков слов у мужчин (1) и женщин (2).
Воспроизведение слов с правого уха не отличалось достоверно у мужчин и женщин. При узнавании у женщин в отличие от мужчин наблюдалась асимметрия воспроизведения, обусловленная лучшим узнаванием слов, предъявлявшихся в левое ухо. У мужчин асимметрия в узнавании латерально предъявленных слов отсутствовала. (Рис.2).
Данные о меньших латеральных различиях показателей непосредственного воспроизведения у женщин по сравнению с мужчинами согласуются с результатами, полученными в аналогичной ситуации дихотического тестирования с применением в качестве стимульного материала серий конкретных существительных (И.А. Вартанян с соавт., 1988). В то же время при использовании в качестве дихотически предъявляемых стимулов названий чисел, одинаковый эффект правого уха выявлен независимо от пола испытуемых (J. McGlone, W. Davidson, 1973; M.P. Bryden, 1975). Если исходить из гипотезы о билатеральном распределении аналогичных речевых функций между полушариями у женщин, то приведенные данные трудно поддаются объяснению, так как в обоих случаях предъявляется вербальная информация.
В отличие от результатов предыдущих исследований, анализировавших показатели асимметрии, рассмотрение нами данных, полученных для каждого полушария, показывает, что различия между мужчинами и женщинами в воспроизведении слов связаны с процессами, происходящими в правом полушарии. Если учесть, что при запоминании конкретных существительных, отличающихся от других вербальных стимулов высокой образностью, могут наряду с левополушарными использоваться правополушарные способы обработки информации, то выявленное правополушарное преимущество у женщин можно объяснить эффективным использованием ими правополушарных стратегий обработки информации. Для проверки этого предположения мы провели изучение стратегий, использовавшихся при запоминании дихотически предъявленных слов. Как видно из рисунка 3 у мужчин доминировала стратегия запечетлевания отдельных слов, а у женщин - образного представления и объединения слов (Н.В. Вольф, 1994, 2000).
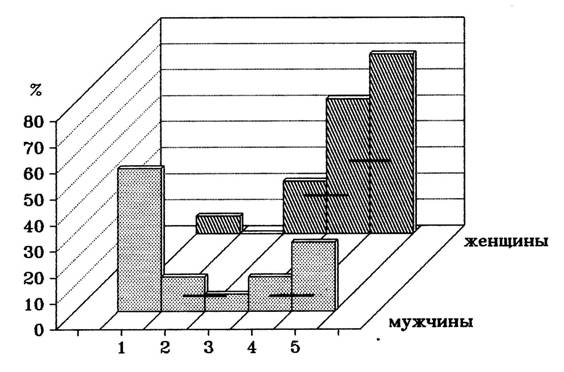
Рис.3. Распределение испытуемых в зависимости от стратегий запоминания слов.
Столбики – процент испытуемых, использующих данную стратегию; горизонтальные линии – количество испытуемых, использующих обе прилегающие стратегии. Стратегии запоминания: 1 – запечатлевание, 2 – по категориям, 3 – по созвучию, 4 – объединение слов в предложение, 5 – образное запоминание.
Анализ стратегий, применявшихся испытуемыми разного пола при запоминании большого объема вербальной информации, позволяет предположить, что преобладающая у мужчин стратегия запечатлевания, для которой характерно "всплывание" слов из памяти при воспроизведении, связана с левополушарными процессами собственно вербальной памяти. Доминирующая у женщин стратегия образного опосредования соответствует правополушарному способу представления и запоминания информации. Показано, что повышение функциональной активности правого полушария при гипнозе активирует образное мышление, стимулирует образную память. При этом по отчетам испытуемых усиливается взаимодействие между образами, что часто сопровождается придумыванием сопутствующих рассказов, чтобы облегчить запоминание (H.J. Crawford, S.N. Allen, 1983; В.С. Ротенберг, 1985). При инактивации левого полушария после применения унилатеральной электросудорожной терапии основным принципом объединения разных слов является их отнесение к внеязыковому предметному миру, воссоздание чувственного наглядного образа (Л.Я. Балонов, В.Л. Деглин, 1976; Л.Я. Балонов c соавт., 1985).
Особого внимания заслуживает тот факт, что наибольшие различия между мужчинами и женщинами обнаружены нами при отсроченном воспроизведении, в ходе которого женщины воспроизводили значительно больше слов, чем мужчины. Существует имеющая подтверждение во множестве экспериментов точка зрения, что способность к воспроизведению из памяти через различные промежутки времени определяется не природой фиксации следа, а возможностью его считывания (Р.Ю. Ильюченок c соавт., 1981). В нашем эксперименте каждый список слов предъявлялся испытуемым однократно. В опытах на животных показано, что определяющую роль в однократном обучении играет эмоциогенная система мозга, способствующая установлению связи следа памяти с программой воспроизведения (Р.Ю. Ильюченок, 1979; Р.Ю. Ильюченок с соав., 1981). При обеспечении процессов вербальной памяти эта регуляторная система, несомненно, работает во взаимодействии с другими, и, прежде всего корковыми, структурами мозга, так как для распознавания эмоциональности слов необходим их семантический анализ. О более тесной связи правого полушария с подкорковыми системами эмоциональной регуляции свидетельствует его преимущественная, по сравнению с левым полушарием, роль в регуляции вегетативных функций (W. Wittling, 1990; B.N. Johnsen, R. Hugdahl, 1993; W. Heller, 1993a). Таким образом, можно предполагать, что выявленное нами у женщин большее вовлечение в процессы вербальной памяти правого полушария, для которого показано тесное взаимодействие с подкорковыми системами эмоциональной регуляции, способствует формированию устойчивых связей следа памяти с программой воспроизведения и высокой эффективности непосредственного и, особенно, отсроченного воспроизведения. Этот вывод подтверждается результатами, полученными нами при изучении влияния эмоциогенной стимуляции (электрокожное раздражение) на воспроизведение дихотически предъявляемых списков слов. Эмоциогенная стимуляция приводила к улучшению отсроченного воспроизведения слов у мужчин и ухудшению – у женщин. Можно полагать, что дополнительная эмоциогенная активация после восприятия информации создает у мужчин оптимальные условия для успешного считывания следа памяти, тогда как у женщин оптимальная активация создается самим мнемическим заданием, и дополнительная эмоциогенная стимуляция в этом случае приводит к ухудшению показателей вербальной деятельности (О.М. Разумникова, Н.В. Вольф, 2002).
Приведенные выше факты касались половых различий в эффективности воспроизведения латерализованной вербальной информации. В то же время известны данные, что процессы, обеспечивающие правильность и скорость мнестических операций могут иметь разную церебральную организацию (R.S. Lewin et al., 1994). В связи с этим для исследований полушарных механизмов памяти представляет интерес модель Стенберга (S. Sternberg, 1966, 1969; R.S. Lewin et al.,1994), позволяющая оценить скорость сканирование памяти. Существует точка зрения, что сканирование памяти в модели Стенберга осуществляется на основе зрительных кодов (S. Sternberg, 1976). Приведенные выше результаты наших исследований указывают на половые различия в использовании правополушарных, образных, способов обработки информации для обеспечения процессов вербальной памяти. В связи с этим представлялось, что исследование скорости сканирования вербальной памяти даст новые представления о полушарной организации вербальных мнестических процессов у мужчин и женщин.
В качестве показателя скорости и правильности сканирования кратковременной памяти анализировали наклон линий (коэффициенты регрессии), отражающих зависимость ВР от объема запоминания. Показатели, полученные при предъявлении цифр в разные полуполя зрения, анализировали отдельно. Как видно из Рис.4, сравнение тестового стимула с удерживаемым в памяти набором быстрее производили женщины.
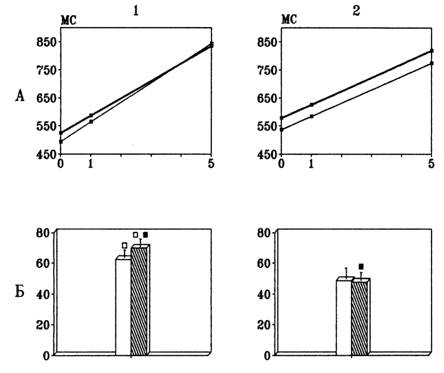
Рис.4. Различия в скорости сканирования кратковременной памяти у мужчин (1) и женщин (2).
А – изменение времени реакции в зависимости от объема запомненной информации: по оси Y- время реакции; по оси X – количество удерживаемых в памяти цифр; толстая линия – правое, тонкое – левое поле зрения. Б – значения коэффициента, характеризующего наклон линии зависимости времени реакции от нагрузки на память. Светлые столбики – правое поле зрения, обозначенные штриховкой – левое;- P<0.05
Хотя тенденция к более быстрой обработке информации наблюдается при предъявлении стимулов как в правое, так и в левое полушарие, только для правого полушария скорость сканирования у женщин была достоверно выше, чем у мужчин. Дальнейший анализ внутригрупповых данных выявил также достоверные латеральные различия коэффициентов, характеризующих наклон линии зависимости ВР от количества элементов в запомненном наборе, у мужчин. Эта зависимость была более выраженной при предъявлении тестовых стимулов в левое полуполе зрения, что может свидетельствовать о разной эффективности мнестических процессов в правом и левом полушариях у мужчин. У женщин наклон линий был одинаков для предъявлений стимулов в правое и левое полуполя зрения.
Возрастание ВР с увеличением набора целевых стимулов, наблюдаемое в наших экспериментах, показывает, что сопоставление пробного стимула с целевым производится последовательно. Принимая во внимание данные о большей эффективности связанных с правым полушарием зрительно-пространственных функций у мужчин, полученные результаты более быстрого сканирования кратковременной памяти женщинами можно объяснить, предположив, что у них более эффективно протекает процесс идентификации зрительных кодов вербальных стимулов. Это предположение хорошо согласуется с нашими данными о том, что использование стратегии образного опосредования при большой скорости предъявления слуховых словесных стимулов является предпочитаемым и эффективным приемом при запоминании вербальной информации женщинами.
Только у мужчин наблюдалась более значительная зависимость ВР от нагрузки на память для правого полушария. Эти данные свидетельствуют о наличии у мужчин асимметрии в воспроизведении данных из кратковременной памяти при меньшей эффективности этого процесса в правом полушарии. Аналогичный результат получен Д. Дж. Маденом и Р. Д.Небесом (D.J. Madden, R.D. Nebes, 1980) при использовании набора цифр на здоровых испытуемых, а также при использовании геометрических стимулов на макаках-самцах (R.S. Lewin et al., 1994). Однако выше были приведены сведения, отличающиеся от полученных нами данных. Наши результаты позволяют заключить, что расхождение данных, обнаруженных в предыдущих исследованиях латерализации процесса сканирования кратковременной памяти, проведенных без учета пола испытуемых, может быть следствием широкого варьирования полового состава экспериментальных групп.
В отличие от мужчин у женщин обнаружено отсутствие различий в наклоне линий, полученных для предъявлений тестирующих стимулов в левое и правое поля зрения, что свидетельствует о более высоких возможностях сканирования вербальной памяти в правом полушарии у женщин.
В исследованиях опознания зрительных образов показано, что описания изображений в правом и левом полушариях не идентичны. В левом полушарии осуществляется инвариантное описание объекта, а в правом - единое полное описание изображения. Соответственно, для опознания образов в левом полушарии используется классификационный, дискриминантный способ, в правом - структурный (Л.И. Леушина с соавт., 1982; А.А. Невская, Л.И. Леушина, 1990). Имея дело с хорошо знакомыми объектами, к которым, в частности, относятся применяемые нами цифровые стимулы, левое полушарие обладает преимуществом в опознании, быстро принимая решение, к какому из известных классов их следует отнести (А.А. Невская, Л.И. Леушина, 1990).
Для оценок полушарного вклада в обработку речевой информации с учетом возможностей образного опосредования большое значение имеет тот факт, что в правом полушарии в процессе тренировки на базе единого полного описания может вторично сформироваться инвариантное описание изображения, аналогичное левополушарному (Л.И. Леушина с соавт., 1982; А.А. Невская, Л.И. Леушина, 1990). Раннее становление речевой функции, широкое использование образного опосредования при осуществлении вербальных функций - все это, на наш взгляд, способствует закреплению за правым полушарием женщин инвариантного описания и дискриминанатного способа обработки зрительной информации, представляющей часто встречающиеся вербальные стимулы. Показано, что тенденция к применению левополушарного дискриминантного способа анализа стимулов проявляется у женщин и при использовании в модели Стенберга невербализируемых пространственных стимулов (С.Б. Цветовский, 1990). Известно, что применение левополушарных стратегий часто оказывается неэффективным при решении сложных зрительно-пространственных задач и, таким образом, может лежать в основе более низких образно - пространственных способностей у женщин по сравнению с мужчинами.
На основании данных о локальном представительстве речевых функций в левом полушарии у женщин и диффузном у мужчин при рассмотрении эффекта более успешного воспроизведения женщинами вербальной информации, адресованной правому полушарию, нами было сделано предположение об ограниченности межполушарных тормозных влияний у женщин и генерализованном тормозном межполушарном взаимодействии у мужчин. Для проверки этого предположения было проведено исследование половых особенностей эффектов интерференционных межполушарных взаимодействий при запоминании вербальной информации. Интерференционные взаимодействия исследованы для 3-х уровней асинхронности предъявления речевых сигналов (-50, 0, +50 мс) на основе сравнения разницы в количестве слов, воспроизведенных соответственно с правого или левого уха при дихотическом (синхронном и асинхронном) и монауральном предъявленьях (Н.В. Вольф, 1998).
Характер интерференционных влияний с правого на левое и с левого на правое полушария представлен на Рис. 5. Как видно из рисунка, латеральные различия интерференционных влияний были выраженными для мужчин и отсутствовали у женщин. У мужчин асимметрия межполушарного интерференционного тормозного влияния характеризовалась его большей его выраженностью с левого на правое полушарие. Известно, что асимметричность тормозных межполушарных влияний способствует "контрастированию" полушарных эффектов и, в соответствии с результатами нашего исследования, может быть механизмом усиления роли левого полушария в речевых процессах у мужчин.
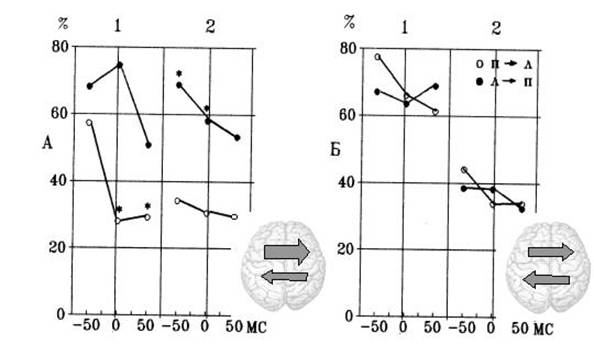
Рис.5. Интерференционное влияние конкурентных вербальных сигналов в зависимости от асинхронности предъявления у мужчин (А) и женщин (Б).
1- первые, 2 – вторые половины списков. По оси Y – интерференционное торможение, по оси X – асинхронность предъявления стимулов в паре: отрицательное значение – запаздывание, положительное – опережение интерферирующего сигнала. * - P<0.05 по сравнению с соответствующим значением в группе женщин.
В свою очередь отсутствие асимметрии межполушарных тормозных влияний может быть следствием локальной организации процессов обработки речевой информации в структурах левого полушария у женщин. Вовлечение в деятельность локальных отделов левого полушария соответствует ограниченности тормозных влияний с левого на правое полушарие у женщин и создает условия для эффективного функционирования негомологичных отделов последнего в вербальных процессах, использования правополушарного образного опосредования при запоминании речевых стимулов, особенно таких, как используемые в наших экспериментах конкретные существительные. В пользу половых различий процессов полушарной интеграции свидетельствуют и наши данные, указывающие на большую скорость межполушарного переноса информации у женщин по сравнению с мужчинами (Н.В. Вольф, О.М. Разумникова, 2002).
Среди электрофизиологических исследований большинство работ, рассматривающих половые различия, связано с анализом ЭЭГ или ВП. Исследования ЭЭГ в ходе выполнения вербального и невербального задания показало снижение мощности альфа-ритма в первом случае в левом, во втором - в правом полушарии в одинаковой степени выраженное у мужчин и женщин (W.H. Moore, 1979). Рассмотрение изменений по сравнению с фоном биопотенциалов альфа- и тета-диапазонов ЭЭГ в каждом полушарии при прослушивании и запоминании словесной информации показало, что у мужчин запечатление слов сопровождалось преимущественной активацией левого полушария и сопутствующим снижением активности правого. Изменения ЭЭГ при запоминании слов у мужчин были значительнее, чем при запечатлении числовой или музыкальной информации, что свидетельствует о наибольшей сложности данного задания. У женщин запоминание словесного материала происходило сравнительно легко, не вызывая заметных ЭЭГ-реакций за исключением снижения мощности альфа-ритма в левой центральной зоне (В.Ф. Коновалов, Н.А. Отмахова, 1984; Н.А. Отмахова, 1987). Анализ данных, полученных в этих исследованиях, позволяет предположить, что их неоднородность может быть связана с разной локализацией отводящих электродов при ограниченном количестве отведений, широким варьированием характера и трудности предъявлявшихся вербальных и пространственных заданий. Так по данным С. Варренбурга и Р. Пагано (S. Warrenburg, R. Pagano, 1981) достоверные половые различия асимметрии ЭЭГ наблюдаются только при выполнении сложных зрительно-пространственных заданий, требующих трехмерного представления. Результаты, свидетельствующие об увеличении сложности задания как факторе, способствующем выявлению половых различий, в частности в электрофизиологических исследованиях, получены и в других исследованиях (А.И. Федотчев, 1985).
В связи с этим при изучении половых различий важное значение имеет выбор адекватного по сложности задания. Однако это условие редко выполняется из-за отсутствия систематических исследований, ставящих целью изучение нейрофизиологической основы половых различий в организации речевых функций. Можно привести лишь единичные работы, в которых изучались особенности активности полушарий у мужчин и женщин при множественной регистрации электрической активности. Так, методом вызванных потенциалов, регистрируемых в 14 симметричных отведениях правого и левого полушарий, показано, что латентные периоды поздних (несенсорных) компонентов вызванных потенциалов на предъявление повторяющихся слов у женщин короче, чем у мужчин. Обнаружены также половые различия вызванных потенциалов, связанные с положением регистрирующих электродов: в вербальной задаче у женщин в задних отделах полушарий амплитуда позитивных компонентов была больше, чем у мужчин; при идентификации повторяющихся фигур, наоборот, для мужчин были характерны большие значения позитивных компонентов в задних отделах полушарий, а для женщин в передних (M.J. Taylor et al., 1990). Анализ изменений ЭЭГ, вызванных сравнением слов, представленных в виде анаграмм, также выявил половые различия в амплитуде и топографии вызванных ответов. Многие различия были обнаружены для коротколатентных (70-120 мс) ответов, что свидетельствует о вовлечении различных корковых ансамблей у мужчин и женщин уже на ранних этапах обработки речевой информации (W. Skrandies et al., 1999). Половые различия в функциональной корковой организации при чтении и построении грамматических конструкций показаны при изучении мозгового кровотока методом позитронной эмиссионной томографии. При генерации глаголов в прошедшем времени у мужчин обнаружено левостороннее, а у женщин билатеральное усиление кровотока в области сильвиевой извилины. В обоих заданиях у женщин была выше активность в затылочных областях мозга (J.J. Jaeger, 1998). При выполнении задания по рифмованию слов левосторонняя активация фронтальной коры у мужчин и более диффузная, захватывающая оба полушария, у женщин обнаружена методом функционального магнитного резонанса (B.A. Shaywitz et al., 1995). Исследование тем же методом латерализации вербальных функций при выполнении двух лексико-семантических задач праворукими испытуемыми показало, что у мужчин они связаны преимущественно с левым полушарием, тогда как примерно у половины женщин имеет место левостороннее доминирование, а у остальных – билатеральное представительство (E.M. Vikingstad et al., 2000).
При рассмотрении коэффициентов корреляции в качестве показателей пространственной синхронизации ЭЭГ обнаружено, что в покое и при выполнении 3 разных вербальных заданий показатели глобальной синхронизации, а также усредненные показатели синхронизации отдельно для каждого полушария и уровень межполушарной синхронизации были выше у мужчин по сравнению с женщинами (Д.П. Белов с соавт., 1997). Однако в большинстве работ выявлена более высокая степень синхронизации электрической активности мозга у женщин. Так, у женщин обнаружена большая межполушарная когерентность биопотенциалов альфа-диапазона ЭЭГ по сравнению с мужчинами при выполнении вербальных заданий (G.J. Beamont et al., 1978), а также в состоянии спокойного бодрствования (M. Corsi-Cabrera et al., 1989; 1993; F.H. Duffy et al., 1996). С этими результатами согласуются данные работы по изучению мозгового кровотока, которые выявили также большее взаимодействие полушарий при запоминании речевой информации у женщин по сравнению с мужчинами (F.B. Wood et al., 1991). Интересные факты получены при изучении корреляций между изменениями метаболизма глюкозы в разных отделах полушарий в покое у мужчин и женщин. У мужчин наблюдалось больше позитивных внутриполушарных, а у женщин - межполушарных корреляций. Кроме этого, у мужчин обнаружены положительные корреляции между показателями метаболической и функциональной асимметрии, то есть высокие показатели фоновой метаболической активности правого полушария сочетались с лучшим выполнением тестов, ориентированных на правополушарные функции. Наоборот, у женщин преобладали отрицательные корреляции, когда более высокий уровень метаболизма в одном полушарии коррелировал с лучшим выполнением тестов, отражающих активность противоположного полушария (N.P. Azari et al., 1995). Из приведенных данных, видно, что изучение функциональных полушарных взаимодействий является одним из наиболее перспективных направлений в исследования полового диморфизма в организации вербальных функций. Общепринятым показателем внутрикорковых взаимодействий считается когерентность биопотенциалов (М.Н. Ливанов, 1972; О.М. Гриндель, 1980; P. Rappelsberger, H. Petsche, 1988). Малочисленность и противоречивость данных о половых различиях в полушарных взаимодействиях при осуществлении вербальных функций побудило нас провести исследование когерентных связей при запоминании дихотически предъявляемой вербальной информации у мужчин и женщин.
Как и в поведенческих исследованиях, мы предъявляли для запоминания семантически значимые вербальные стимулы (списки конкретных существительных), теоретически допускающих использование как левополушарных, так и правополушарных способов обработки информации. По нашему мнению, именно такие условия должны были способствовать проявлению внутренне присущих испытуемым разного пола полушарных стратегий обработки речевой информации и их нейрофизиологических коррелятов.
При топографическом анализе изменений синхронизации биоэлектрических колебаний обнаружено что запоминание дихотически предъявляемых списков слов сопровождается повышением внутриполушарной когерентности ЭЭГ в тета- и альфа-диапазонах преимущественно в левом полушарии у мужчин и в правом у женщин (Рис.6).
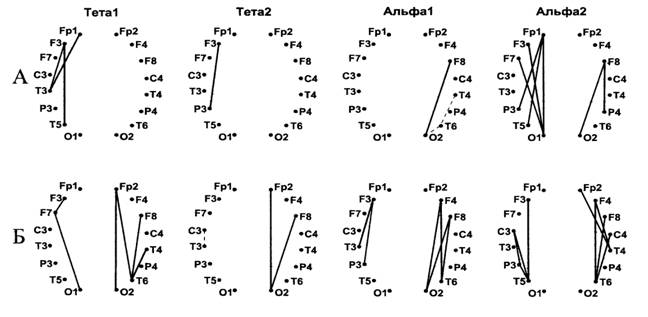
Рис.6. Карты изменений внутриполушарной когерентности у мужчин (А) и женщин (Б) при запоминании дихотически предъявленных списков слов.
Достоверные (P<0.05 по критерию Вилкоксона) изменения когерентности между двумя отведениями представлены соединяющими их линиями. Сплошные линии – возрастание, пунктирные – снижение когерентности по сравнению с фоном.
При этом обращает на себя внимание тот факт, что половые различия касались в основном правого полушария. В тета-диапазоне при преимущественном росте когерентности биопотенциалов в правом полушарии у женщин рост когерентности биопотенциалов в правом полушарии у мужчин вообще отсутствовал. В альфа-диапазоне у женщин в правом полушарии количество пар отведений, для которых отмечен рост когерентности при выполнении вербального задания, более, чем в два раза, превышало количество таких пар у мужчин. Эти результаты, с учетом данных о функциональной значимости параметра когерентности, могут свидетельствовать о более высокой функциональной активности правого полушария в процессе выполнения вербального задания у женщин по сравнению с мужчинами.
Половой диморфизм обнаружен и при анализе изменений когерентности между отведениями, относящимися к разным полушариям. В тета диапазонах у женщин в отличие от мужчин наблюдалось более значительное и диффузное увеличение межполушарной когерентности. Качественные различия функциональной полушарной организации у мужчин и женщин проявились в пространственном распределении измененных межполушарных когерентных связей при запоминании речевой информации. У женщин наблюдалось усиление связей преимущественно передних ассоциативных областей коры каждого полушария с пространственно удаленными задними отделами коры противоположных полушарий. У мужчин изменения межполушарной когерентности ограничивались парами отведений, представлявших задние отделы правого и левого полушарий (Рис.7). При этом у женщин отмечены преимущественно положительные корреляции между эффективностью воспроизведения и возрастанием межполушарной когерентности.
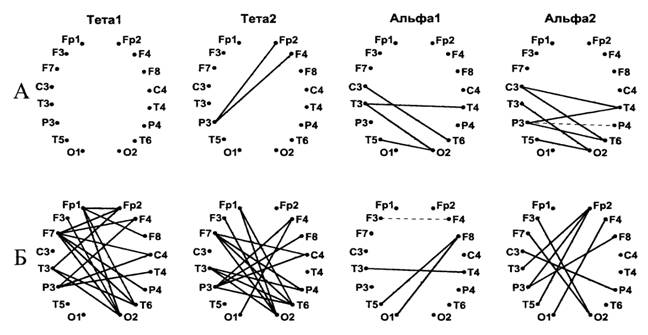
Рис.7. Карты изменений межполушарной когерентности у мужчин (А) и женщин (Б) при запоминании дихотически предъявленных списков слов.
Обозначения как на рис.6.
У мужчин коэффициенты корреляции между соответствующими показателями имели отрицательные значения (Рис.8).
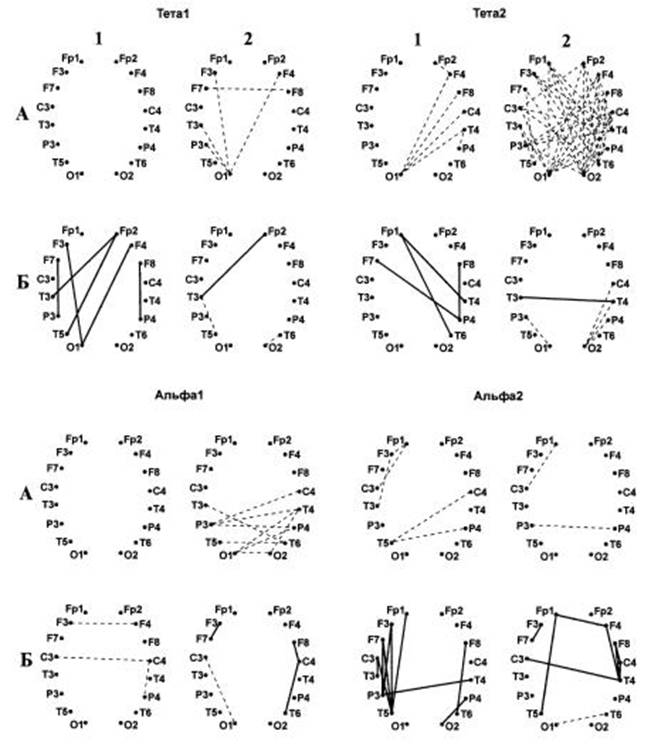
Рис.8. Карты изменений когерентности биопотенциалов, коррелирующих с эффективностью воспроизведения слов, адресованных левому (1) и правому (2) полушариям.
А – мужчины, Б- женщины. Сплошные линии – достоверные (r>0.6, P<0.05) положительные корреляции, пунктирные – отрицательные.
Известно, что ассоциативные зоны лобных отделов мозга играют ведущую роль в организации активного мнестического процесса, мало влияя на пассивное запечатление материала (А.Р. Лурия, 1973). Усиление функциональных связей между фронтальными и сложными межанализаторными каудальными ассоциативными зонами противоположных полушарий может создавать условия для кооперации фронтальной коры с височными областями, участвующими в анализе устной речи, и парието-окципитальными отделами, обеспечивающими обработку зрительных образов (P. Rappelsberger, H. Petche,1995). В связи с этим стоит напомнить приведенные ранее данные об использовании большинством женщин стратегии образного опосредования при запоминании дихотически предъявляемых существительных. Основываясь на данных настоящего исследования, можно предположить, что женщины более успешно запоминают конкретные существительных за счет включения полимодальных сенсорных стратегий, базирующихся на кооперации функционально различных отделов мозга.
В пользу этого свидетельствуют и данные, полученные нами при исследовании ЭЭГ в процессе воспроизведения дихотически предъявленных списков слов. При анализе изменений мощности ЭЭГ, вызванных воспроизведением, половые различия выявлены для биопотенциалов тета2 диапазона. У женщин при воспроизведении наблюдалось более выраженным, чем у мужчин снижение мощности ритма в парието-окципитальных и задневисочных отделах коры мозга. Существуют данные, что мысленное воспроизведение прослушанных слов и связанных с ними образов приводит к активации, как слуховой коры, так и парието-окципитальных областей, связанных со зрительно-пространственными представлениями, пространственным воображением и вниманием (S.P. Gandhi, 2001). В соответствии с этими данными можно предположить, что рассмотренные выше половые особенности кодирования запоминаемой информации накладывают отпечаток на паттерны активности мозга при воспроизведении, что отражается в большей десинхронизации тета2-ритма в задних отделах коры мозга у женщин по сравнению с мужчинами.
В тета2, альфа1 и альфа2 частотных диапазонах у женщин наблюдались достоверные латеральные различия реактивности когерентности, обусловленные возрастанием при воспроизведении когерентности в левом и снижением в правом полушариях. У мужчин латеральные различия отсутствовали (Рис.9).
Существуют модели эпизодической памяти, в которых предполагается, что воспроизведение приводит к восстановлению паттерна нервной активности, свойственного стадии фиксации информации (A.R. Damasio, 1989; L.R. Squire, P. Alvarez, 1995). Однако в ряде исследований такое соответствие не подтверждается (E. Tulving et al., 1994; K. Allan, W.G.K. Robb, M.D. Rugg, 2000) Наши результаты также указывают на неидентичность нервных процессов, лежащих в основе запоминания и воспроизведения следа памяти. Для всех рассматриваемых частотных диапазонов у женщин латеральные различия в реактивности когерентности были обусловлены снижением когерентных взаимодействий в правом и повышением в левом полушарии в ходе воспроизведения. В томографических исследованиях показано, что воспроизведение из семантической памяти в большей степени связано с левым, а из эпизодической – с правым полушарием (E. Tulving et al., 1994; R.L. Buckner, E. Tulving, 1995).
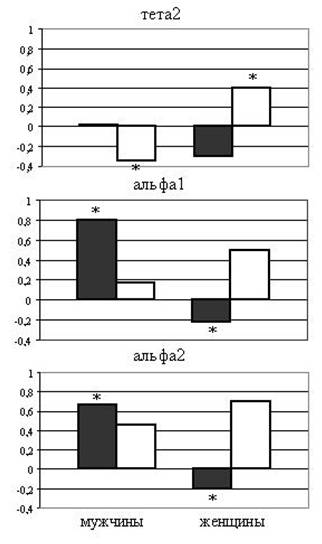
Рис.9. Латеральные различия связанной с воспроизведением реактивности когерентности (разности между значениями в фоне и при воспроизведении) у мужчин и женщин
Темные столбики - суммарная реактивность когерентности левого полушария, светлые - правого. * - P<0,05 между одинаково обозначенными значениями в данном частотном диапазоне.
Таким образом, сдвиг баланса внутрикорковых взаимодействий в сторону левого полушария согласуется с нашими данными о более глубокой семантической переработке запоминаемой информации у женщин, что приводит к преобладанию семантического воспроизведения над эпизодическим. Напротив, большие значения когерентности тета2 ритма в правом полушарии у мужчин по сравнению с женщинами могут свидетельствовать о доминировании у них воспроизведения из эпизодической памяти, для которого необходима постоянная связь воспроизведения с маркерами предшествующего эпизода запоминания.
В наших экспериментах выявлены достоверные половые различия в реактивности когерентности биопотенциалов альфа1 и 2 диапазонов в левом полушарии, которые были обусловлены усилением когерентных взаимодействий при воспроизведении у женщин и снижением у мужчин. Что касается альфа1 ритма, то изменения когерентности биопотенциалов этого частотного диапазона позволяют дифференцировать обработку информации различной сенсорной модальности, а изменения альфа2 ритма отражают особенности семантической обработки информации (S.Weiss, P. Rappeisberger, 2000). Приведенные данные позволяют заключить, что у женщин воспроизведение конкретных существительных обеспечивается за счет формирования в левом полушарии динамических функциональных систем, возможно, связанных с процессами сознательной реконструкции сенсорных и семантических характеристик запомненной информации.
Половые различия ЭЭГ коррелятов процессов фиксации и воспроизведения слов могут быть связаны с упоминавшимися ранее особенностями представительства речевых функций в левом полушарии у мужчин и женщин (D. Kimura 1980; 1983; C. Mateer et al., 1982; G.A. Ojemann 1983; G.A. Ojemann et al., 1989), которые могут создавать условия для параллельного участия обоих полушарий и их взаимодействия в процессе обработки вербальной информации у женщин. В свою очередь выявленные нами при запоминании слов множественные отрицательные корреляции между эффективностью воспроизведения информации, адресованной правому полушарию, и усилением межполушарных связей у мужчин могут свидетельствовать о тормозном характере межполушарных взаимодействий.
Приведенные данные в совокупности с рассмотренными ниже современными представлениями о механизмах межполушарных взаимодействий позволяют предложить новую гипотезу относительно половых различий в организации полушарных функций.
Что касается межполушарных взаимодействий, то наибольшей экспериментальной и теоретической разработке подверглись представления о тормозных взаимодействиях и роли мозолистого тела в поддержании процессов независимой обработки информации в каждом из полушарий, "экранировании" деятельности одного полушария от другого (V.H. Deneberg, 1983; N.D. Cook, 1986; E. Zaidel et al., 1990; T.V. Wey et al., 1993; C. Chiarello, L. Maxfield, 1996). Часто рассматривают одностороннее торможение одного полушария другим. О торможении правого полушария со стороны левого обычно говорят в связи с речью. В пользу тормозного влияния левого полушария на правое свидетельствуют факты повышения речевой компетенции правого полушария у лиц, перенесших операцию расщепления мозга (M.S. Gazzaniga, R.W. Sperry, 1967; M.S. Gazzaniga, S.A. Hillard, 1971). Снижение функциональных возможностей левого полушария, связанное с наличием подкорковых фокусов судорожной активности, также ведет к расширению возможностей правого полушария в выполнении несвойственных ему речевых функций (M. Regard et al., 1994). С точки зрения однонаправленного торможения левым полушарием правого объясняется факт снижения амплитуды вызванных потенциалов на зрительные стимулы в левом полушарии и повышения в правом в процессе решения испытуемыми вербальных задач. Предполагается, что повышение активации левого полушария во время обработки речевого материала (обнаружение слов одной семантической категории или фонетический анализ) приводит к уменьшению амплитуды, регистрируемых в этом полушарии вызванных потенциалов на зрительные стимулы. Увеличение амплитуды вызванных потенциалов в правом полушарии является отражением снижения его функциональной активности и активации. На основе анализа полученных данных автор делает вывод об активном торможении левым полушарием гомологичных отделов правого полушария в процессе обработки речевых сигналов (A.C. Papanicolau, 1980).
Последние модели тормозного межполушарного взаимодействия отходят от представлений о глобальном тормозном влиянии одного полушария на другое, заменяя их моделями ограниченных межполушарных взаимодействий (N.D.Сook, 1986; T.V. Wey et al., 1993). Основываясь на данных нейроанатомии и нейрофизиологии мозолистого тела, Н.Д. Кук (N.D. Cook, 1984, 1986) рассматривает следующий механизм тормозных внутри- и межполушарных взаимодействий при обработке речевой информации. Активация речевых зон левого полушария приводит к ипсилатеральному торможению окружающих нейронов и транскаллозальному торможению симметричных отделов правого полушария, сопровождающемуся активацией окружающих нейронов, которые, предположительно, могут участвовать в контекстуальном анализе (W. Wapner et al., 1981). Поставленные ими (T.V. Wey et al., 1993) специальные эксперименты с регистрацией успешности идентификации слов, предъявляемых тахистоскопически при различных вариантах стимуляции контрлатерального зрительного поля, подтвердили положения, предсказываемые предлагаемой моделью межполушарного торможения.
Т.В.Уэй с соавторами (N.V. Wey et al., 1993) предполагают, что сила тормозных влияний каждого из полушарий пропорциональна степени его вовлечения в конкретную деятельность. Другими словами, полушарие, доминирующее в данном задании, будет оказывать более выраженный ингибирующий эффект на противоположное полушарие, усиливая межполушарные различия. При этом преимущественно гомотопный характер межполушарных связей будет способствовать подавлению только идентичных процессов в противоположном полушарии.
Идея локальных межполушарных взаимодействий допускает одновременное участие двух полушарий, осуществляющих специфические функции, в решении любой задачи. Таким образом, результаты межполушарного взаимодействия могут зависеть как от характера вербальных сигналов и модальности их предъявления, так и от специфики задания, определяющих возможности использования и соотношение левополушарных и правополушарных стратегий обработки информации.
Таким образом, диффузное представительство речевых функций в левом полушарии у мужчин может приводить к диффузному торможению правого полушария при осуществлении вербальных мнестических функций. В отличие от мужчин у женщин тормозные влияния с левого полушария на правое могут быть менее генерализованными в связи с фокальным представительством речевых функций в их левом полушарии. При этом локальность тормозных влияний может способствовать кооперации правого и левого полушарий для обеспечения вербальных функций у женщин при значительном участии корково-подкорковых связей. Рисунок 10 является графическим представлением предложенной модели.
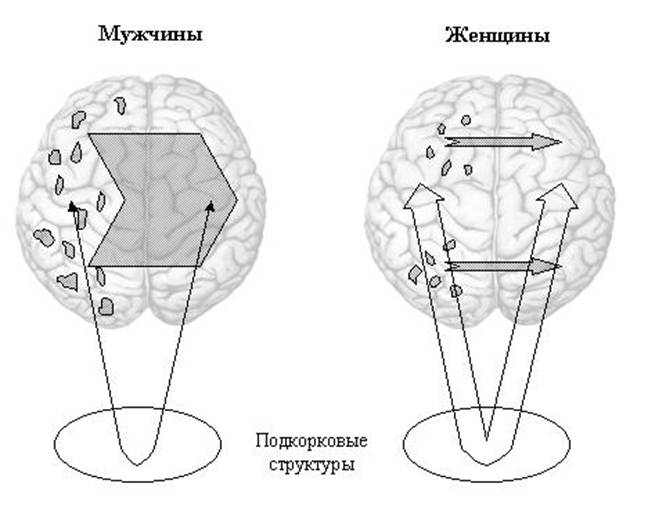
Рис.10. Модель межполушарных и корково-подкорковых взаимодействий при осуществлении вербальных функций у мужчин и женщин.
Серые области в левом полушарии – зоны представительства вербальных функций. Тонированные стрелки – гомотопное трансколлозальное торможение с левого на правое полушарие; незакрашенные - межполушарные взаимодействия через подкорковые структуры.
ЛИТЕРАТУРА
Балонов Л.Я., Деглин В.Л. Слух и речь доминантного и недоминантного полушарий. - Л.: Наука, 1976. - 220 С.
Балонов Л. Я., Деглин В.Л., Черниговская Т.В. Функциональная асимметрия мозга в организации речевой деятельности // Сенсорные системы: Сенсорные процессы и асимметрия полушарий. - Л.: Наука, 1985. - С.99-115.
Белов Д.П. Кануников М.Е. Кавшбая Н.А. Влияние пола на пространственную синхронизацию ЭЭГ // Российск. физиол. журн. – 1997. – Т.83. – N7. – С.28-36.
Вартанян И.А., Галунов В.И., Дмитриева Е.С., Зайцева К.А., Королева И.В., Кузьмин Ю.И., Морозов В.П., Шургая Г.Г. Восприятие речи. Вопросы функциональной асимметрии мозга. - Л.: Наука, 1988. - 135 C.
Вольф Н.В. Половые различия при запоминании дихотически предъявленных списков слов // Журн. высш. нервн. деят. - 1994. - Т.44. - Вып.1. - С.18-24.
Вольф Н.В. Половые различия межполушарных интерференционных взаимодействий при запоминании речевой информации // Журн. высш. нервн. деят. - 1998. - Т.48. - N3. - С.551-553.
Вольф Н.В.,. Разумникова О.М. Время унимануальных реакций при сравнении латерализованных вербальных стимулов: особенности межполушарных взаимодействий, связанные с полом испытуемых // Физиол. челов. - 2002. – Т.28. – N4. - С.
Вольф Н.В.,. Разумникова О.М. Половые различия полушарных пространственно-временных паттернов ЭЭГ при воспроизведении вербальной информации // Физиол. челов. - 2002. – Т.28. – N6. - С.
Гриндель О.М. Оптимальный уровень когерентности ЭЭГ и его значение в оценке функционального состояния мозга человека // Журн. высш. нервн. деят. - 1980. - Т.30. - Вып.1. - С.62-70.
Ильюченок Р.Ю. Память и адаптация. - Новосибирск: Наука, 1979. - 192 С.
Ильюченок Р.Ю., Гилинский М.А., Лоскутова Л.В., Дубровина Н.И., Вольф Н.В. Миндалевидный комплекс. - Новосибирск: Наука, 1981. - 242 С.
Кауфман Д.А., Траченко О.П. Функциональная асимметрия мозга и восприятие слов, относящихся к разным грамматическим категориям // Сенсорные системы: Сенсорные процессы и асимметрия полушарий.- Л.: Наука, 1985.- С.115-126.
Коновалов В.Ф., Отмахова Н.А. Межполушарная асимметрия ЭЭГ-реакций // Психол. журн. - 1984. - Т.5. - N1. - С.62-70.
Леушина Л.И., Невская А.А., Павловская М.Б., Вершинина Е.А. Инвариантность зрительного опознания в правом и левом полушариях мозга человека // Переработка информации в зрительной системе. Высшие зрительные функции. - Л.: Наука, 1982. - С.21-36.
Ливанов М.Н. Пространственная организация процессов головного мозга.- М.: Наука, 1972. - 182 С.
Лурия А.Р. Основы нейропсихологии.-М.:Изд. МГУ, 1973.- 388 С.
Невская А. А., Леушина Л. И. Асимметрия полушарий и опознание зрительных образов. - Л.: Наука, 1990. - 152 С.
Отмахова Н.А. Половые особенности межполушарной асимметрии // Асимметрия мозга и память. - Пущино, 1987. - C.115-124.
Разумникова О.М., Вольф Н.В. Половые различия временной динамики межполушарной асимметрии восприятия речевой информации // Бюл. Сибирск. Отдел. РАМН - 1997. - N2. - C.87-90.
Разумникова О.М., Вольф Н.В. Латеральные эффекты эмоциогенной стимуляции в вербальных мнестических процессах у мужчин и женщин // Журн. высш. нервн. деят. - 2002. - Т.52. - Вып.3. – С197-192.
Ротенберг В.С. Гипноз и образное мышление // Психологич. журн. - 1985. - Т.6. - N2. - C.128-133.
Спрингер С., Дейч Г. Левый мозг, правый мозг. - М.: Мир. - 1983. - 256 С.
Трубникова Р.С. ЭЭГ, память и половой диморфизм // Память и следовые процессы. - Пущино, 1974. - С.248-249
Федотчев А.И. Проявление половых различий в психофизиологических показателях в зависимости от сложности экспериментального задания // Физиол. челов. - 1985. - Т.11. - N5. - С.730-733..
Цветовский С.Б. Специализация полушарий и индивидуальные особенности межполушарной функциональной асимметрии в процессах опознания зрительной информации // Бюлл. Сибирск. Отдел. РАМН. - 1992. - N1. - С.38-44.
Черниговская Т.В., Лях Н.Ю., Токарева Т.И. Латерализация восприятия слов в зависимости от лингвистических факторов // Физиол. челов. - 1996. - Т.22. - N4. - C.12-17.
Aboitiz F., Sheibel A.B., Fisher R.S., Zaidel E. Fiber composition of the fuman corpus callosum // Brain Behav. Evol. - 1992. - Vol.598. - P.143-153.
Aboitiz F., Sheibel A.B., Zaidel E. Morphometry of the Sylvian fissure and the corpus callosum, with emphasis on sex differences // Brain. - 1992. - Vol.115. - P.1521-1541.
Allan K., Robb W.G.K., Rugg M.D. The effect of encoding manipulations on neural correlates of episodic retrieval // Neuropsychologia. - 2000. - Vol.38 - N8. P.1188- 1192..
Azari N.P., Pettigrew K.D., Pietrini P., Murphy D.G., Horwitz B., Shapiro M.B. Sex differences in patterns of hemispheric cerebral metabolism: a multiple regression/discriminant analysis of positron emission tomographic data // Intern. J. Neurosci. - 1995. - Vol.81. - N1-2. - P.1-20.
Batler S. Sex difference in human cerebral function // Prog. Brain Res. - 1984. - Vol.61. - P.132-144.
Beamont G.J., Mayers A.R., Rugg M.D. Asymmetry in EEG alfa coherence and power: effects of task and sex // EEG and Clin Neurophysiol. - 1978. - Vol.45. - N3. - P.393-401.
Bleeker M.L., Bolla-Wilson K., Agnew J., Meyers D.A. Age-related sex differences in verbal memory // J. Clin. Psychol. - 1988. - Vol.44. - P.403-411.
Bolla-Wilson K., Bleeker M.L. Influence of verbal intelligence , sex,age,and education on the Rey Auditory Verbal Learning Test // J. Exper.Psychol.:General. - 1986. - Vol.2. - P.203-211.
Bryden M.P. Perceptual asymmetry in vision: Relation to handedness, eyedness, and speech lateralization // Cortex. - 1973. - Vol.9. - P.419-435.
Bryden M.P. Speech lateralization in families: a preliminary study using dichotic listening // Brain Lang. - 1975. - Vol.2. - P.201-211.
Bryden M.P. An overview of the dichotic listening procedure and its relation to cerebral organization // Handbook of Dichotic Listening. - Chichester, England: Wiley, 1988. - P.1-43.
Bryden M.P., Hecaen H., DeAgostini M. Patterns of cerebral organization // Brain Lang. - 1983. - Vol.20. - P.249-262.
Bryden M.P., Munhall K., Allard F. Attentional biases and the right-ear effect in dichotic listening // Brain Lang. - 1983. - Vol.18. - P. 236-248.
Buckner R.L., Tulving E. Neuroimaging studies of memory: theory and recent PET results // Handbook of Neuropsychology. - 1995. - Vol.10. - P.439- 452.
Byne W., Bleier R., Houston L. Variations in human corpus callosum do not predict gender: a study using magnetic resonance imaging // Behav. Neurosci. - 1988. - Vol.102. - P.222-227.
Chiarello C., Maxfielf L. Varieties of interhemispheric inhibition, or how to keep a good hemisphere down // Brain Cogn. - 1996. - Vol.30. - P.81-108.
Clarke S., Kraftsik R., Van der Loos H., Innocentti G.M. Forms and measures of adult and developing human corpus callosum: Is there sex dimorphism? // J. Comp. Neurol. - 1989. - Vol.280. - P.213-230.
Clarke J. M., Zaidel E. Anatomical-behavioral relationships: corpus callosum morphometry and hemispheric specialization // Behav. Brain Res. - 1994. - Vol.64. - P.185-202.
Cook N.D. Callosal inhibition: the key to the brain code // Behav. Sci. - 1984. - Vol.29. - N1. - P.98-110.
Cook N.D. The brain code: mechanisms of information transfer and the role of the corpus callosum. - London: Mathuen, 1986. - 227 P.
Corsi-Cabrera M., Herrera P., Malvido M. Correlation between EEG and cognitive abilities: sex differences .// Int. J..Neurosci . – 1989. – Vol.45. - N1-2. – P.133-141.
Corsi-Cabrera M., Ramos J., Guevara M.A., Arce C., Gutierrez S. Genin the EEG during cognitive activity .// Int. J. Neurosci. – 1993. – Vol.72. - N3-4. – P.257-264.
Crawford H.J., Allen S.N. Enhanced visual memory during hypnosis as mediated by hypnotic responsiveness and cognitive strategies // J. Exp. Psychol. Gen. - 1983. - Vol.112. - N4. - P.662-685.
Damasio A.R. Time-locked multiregional retroactivation: a systems-level proposal for the neural substrates of recall and recognition// Cognition. - 1989. - Vol.33.- N1-2.- P.25 – 31.
Day J. Visual half-field word recognition as a function of syntactic class and imagability // Neuropsychologia. - 1979. - Vol.17. - P.515-523.
Denenberg, V.H. Micro and macro theories of the brain // Behav. Brain Sci. - 1983. - Vol.6. - P.174-178.
Denenberg, V.H., Kertesz A., Cowell P.E. A factor analysis of the human's corpus callosum // Brain Res. - 1991. - Vol.548. - P.126-132.
Duffy F.H., Mcanulty G.B., Albert M.S. Effects of age upon interhemispheric EEG coherence in normal adults // Neurobiol. Aging. - 1996. - Vol.17. - N4. - P.587-99.
Ellis H.D., Shepherd J. W. Recognition of abstract and concrete words presented in left and right fields // J. Exper. Psychol. - 1974. - Vol.103. - P.1035 - 1036.
Ely P.W., Graves R.F., Potter S.M. Dichotic listening indices of right hemisphere semantic prosessing // Neuropsychol. - 1989. - Vol.27. - P.1007-1015.
Foundas A.L., Faulhaber J.R., Kulynych J.J., Browning C.A., Weinberger D.R. Hemispheric and sex-linked differences in Sylvian fissure morphology: a quantitative approach using volumetric magnetic resonance imaging. //Neuropsychiatry Neuropsychol Behav Neurol. – 1999. – Vol. 12. - P. 1-10.
Gandhi S.P. Memory retrieval: Reactivating cortex // Current Biol. - 2001. - Vol.11. - R.32
Gazzaniga M.S., Hillgard S.A. Language and speech capacity of the right hemisphere // Neuropsychologia. - 1971. - Vol.9. - P.273-280.
Gazzaniga M.S., Sperry R.W. Language after section of the cerebral commissure // Brain. - 1967. - Vol.90. - P.131-148.
Geffer G., Moar K.J., O'Hanlon A.P., Clark C.R., Geffen L.B. Performance measures of 16-to-86-years-old males and females on the Auditory Verbal Learning Test //Clin. Neuropsychologist. - 1990. - Vol. 4. - P.45-63.
Gur R.C., Turetsky B.I., Matsui M., Yan M., Bilker W., Hughett P., Gur R.E. Sex differences in brain gray and white matter in healthy young adults: correlations with cognitive performance // J. Neurosci. - 1999. - Vol.9. - N10. - P.4065-4072.
Habib M., Gayraud D., Oliva A., Regis J., Salamon G., Khalil R. Effects of handedness and sex on the morphology of the corpus callosum: a study with brain magnetic resonance imaging // Brain Cogn. - 1991. - Vol.16. - P.41-61.
Harasty J., Double K.L., Halliday G.M., Kril J.J., McRitchie D.A. Language-associated cortical regions are proportionally larger in the female brain. // Arch Neurol. - 1997. – Vol. 54. - N2. – P. 171-176.
Heller W. Neuropsychological mechanisms of individual differences in emotion, personality, and arousal // Neuropsychol. - 1993a. - Vol.7. - N4. - P.476-489.
Herlitz A., Nilson L.-G., Backman L. Gender differences in episodic memory // Memory Cogn. -1997. - Vol.25. - N6. - P.801-811.
Herrman D.J., Crawford M., Holdsworth M. Gender-linked differences in everyday memory performance // Brit. J. Psychol. - 1992. - Vol.83. - N2. - P.221-231.
Hines M., Chiu L., McAdams L.A., Bentler P.M., Lipcamon J. Cognition and the corpus callosum: verbal fluency, visuospatial ability, and language lateralisation related to midsagittal surface areas of callosal subregions // Behav. Neurosci. - 1992. -Vol.106. - N1. - P.3-14.
Hill R.D., Grut M., Wahlin A., Herlitz A., Winblad B., Backman L. Predicting memory performance in optimally healthy very old adults // J. Ment. Health Aging. - 1995. - Vol.1.- P.55-65.
Hiscock M., MacKay M. The sex difference in dichotic listening: multiple negative findings // Neuropsychologia. - 1985. - Vol.23. - P.437-440.
Holloway R.L., Fnderson P.J., Defendini R., Harper C. Sexual dimorphism in the human corpus callosum from three independent samples: relative size of the corpus collosum, Am. J. // Phys Anthropol. -1993. - Vol. 92 - P.481-498.
Holloway R. L., de Lacoste M. C. Sexual dimorphism in the human corpus callosum: An extension and replication study. // Human Neurobiology – 1986. - Vol. 5. - P. 87-91.
Hultsch D. F., Masson M.E.J., Small B.J. Adult age differences in direct and indirect tests of memory // J. Geront.: Psychol. Sci. - 1991. - Vol.46. - P.22-30.
Jaeger J.J., Lockwood A.H., Van Valin R.D.Jr., Kemmerer D.L., Murphy B.W., Wack D.S. Sex differences in brain regions activated by grammatical and reading tasks // Neurorep. - 1998. - Vol.9. - N12. - P.2803-2807.
Johnsen B.N., Hugdhal R. Right hemisphere representation of autonomic conditioning to facial emotional expressions // Psychophysiol. -1993. -Vol.30. -P.274-278.
Jones E.B., Gorman J.G., Byrne B. Forgetting of word associates as a function of recall interval // Brit. J. Psychol. - 1987. - Vol.78. - P.79-89.
Kelly R., Orton K. Dichotic perception of word-pairs with mixed image values // Neuropsychol. - 1979. - Vol.17. - N3-4. - P.363-372.
Kertesz A., Polk M., Black S. E. and Howell J. Sex, handedness, and the morphometry of cerebral asymmetries on magnetic resonance imaging // Brain Res. -1990. - Vol. 530. – P. 40-48.
Kimura D., Sex differences in intrahemispheric organization of speech // Behav.. Brain Sci. - 1980. - Vol.3. - P.240-241.
Kimura D. Sex differences in cerebral organization for speech and praxis functions // Can. J. Psychol. - 1983. - Vol.37. - P. 19-35.3
Kimura D., Harshman R.A. Sex differences in brain organization for verbal and non-verbal function // Prog. Brain Res. - 1984. - Vol.61. - P.423-441.
Kinsbourne M. The control of attention by interaction between the cerebral hemispheres. In: S. Kornblum (Ed.), Attention and Performance IV.// Academic press, New York. - 1973. - P.239-256.
Kulynych J.J., Vladar K., Jones D.W., Weinberger D.R. Gender differences in the normal lateralization of the supratemporal cortex: MRI surface-rendering morphometry of Herschl's gyrus and planum temporale // Cereb. Cortex. - 1994. - Vol.4. - P.107-118.
Lake D.A., Bryden M.P., Handedness and sex differences in hemispheric asymmetry // Brain Lang. - 1976. - Vol. 3. - P. 266-282.
Larrabee G.J., Crook T.H. Do men show more rapid age-associated decline in simulated everyday verbal memory than do women? // Psychol. Aging. - 1993. - Vol. 8. - P.68-71.
Levy J. Cerebral asymmetry and the psychology of man // Brain and Psychology. - N.Y., 1980. - P.183-191.
Levy J., Heller W. Gender differences in human neuropsychological function // Sexual Differentiation, Handbook Behav. Neurol. - N.Y.:Plenum Press, 1992. - Vol.11. - P.245-274.
Lewine J., Doty R., Astur R., Provencal S. Role of the forebrain commissures in bihemispheric mnemonic integration in macaques // J. Neurosci. - 1994. - Vol.14. - N5. - P.2515 - 2530.
Lewis R.S., Christiansen L. Intrahemispheric sex differences in the functional representation of language and praxic functions in normal individuals // Brain Cogn. - 1989. - Vol.9. - P.238-243.
Madden D.J., Nebes R.D. Hemispheric differences in memory search // Neuropsychologia. - 1980. - Vol.18. - P.665 - 673.
Mateer C., Polen S.B., Ojemann G.A. Sexual variation in cortical localization of naming as determined by stimulation mapping // Behav. Brain Sci. - 1982. - Vol.5. - P.310-311.
McGlone J., Sex differences in functonal brain asymmetry // Cortex. - 1978. - Vol.14. - P.122-128.
McGlone J. Sex differences in human brain asymmetry: a critical survey // Behavior. Brain Sci. - 1980. - Vol.3. - P.215-227.
McGlone J., Davidson W., The relationship between cerebral speech laterality and spacial ability with spetial reference to sex and hand preference // Neuropsychologia. - 1973. - Vol.11. - P.105-113.
Mirabile P. J., Porter R. J., Hughes L.F., Berlin Ch. I. Dichotic LAG effect in children 7 to 15 // Devel. Psychol. - 1977. - P.2-8.
Moore W.H. Jr. Alpha hemispheric asymmetry of males and females on verbal and non-verbal tasks: some preliminary results // Cortex. - 1979. - Vol.15. - N2. - P.321-326.
Munro P., Govier E., Dynamic gender-related differences in dichotic listering performance // Neuropsychologia. - 1993. - Vol.31. - N4. - P.347-353.
Ojemann G.A. Brain organization for language from the perspective of electrical stimulation mapping // Behav. Brain Sci. - 1983. - Vol.6. - P.189-230.
Ojemann G.A., Ojemann 0. J., Lettich E., Berger M. Cortical language lateralization in left, dominant hemisphere. An electrical stimulation mapping investigation in 117 patients // J. Neurosurg. - 1989. - Vol.71. - P.316-326.
Orenstein H.B, Meighan W. B. Recognition of bilaterally presented words varying in concreteness and frequency: lateral dominance or sequential processing? // Bull. Psychonom. Soc. - 1976. - Vol.7. - P.179-180.
Papanicolau A.C. Cerebral excitation profiles in language processing: the photic probe paradigm // Brain Lang. - 1980. - Vol.9. - N3. - P.269-280.
Portin R,, Saarijarvi S,, Joukamaa M,, Salokangas R.K. Education, gender and cognitive performance in a 62-year-old normal population: results from the Turva Project // Psychol. Med. - 1995. - Vol.25. - N6. - P.1295-1298.
Prokop A., Oehmichen M., Zilles K. Sexual dimorphism of the corpus callosum? // Beitr. Gerichtle. Med. – 1990. - Vol.48. - P. 263-270.
Rabbitt P., Donlan C., Watson P., McInnes L., Bent N. Unique and interactive effects of depression, age, socioeconomic advantage, and gender on cognitive performance of normal healthy older people // Psychol. Aging. - 1995. - Vol.10. - P.307-313.
Rappelsberger P., Petsche H. Probability mapping: power and coherent analysis of cognitive processes. // Brain Topogr. - 1988. - Vol.1. - P.46-54.
Rappelsberge, P., Petsche H. () Amplitude and coherence analysis of visual imagery and visual perseption. In M. Eiselt, U. Zweiner and H. Witte (Eds.), Quantitative and Topological EEG and MEG Analysis, Universitatsverlag, Jena. 1995, pp.181-186.
Regard M., Cook N.D., Wieser H.G., Landis T., The dinamics of cerebral dominance during unilateral limbic seizures // Brain. - 1994. - Vol.117 .- P.91-104.
Ruff R.M., Light R.H., Quayhagen M. Selective reminding tests: a normative study of verbal learning in adults // J. Clin. Exp. Neuropsychol., 1989. - Vol.11. - P.539-550.
Schaie K.W., Willis S.L. Age difference patterns of psychometric intelligence in adulthood: generalizability withing and across ability domains // Psychol. Aging. - 1993. - Vol.8. - P.44-55.
Schlaepfer T.E., Harris G.J., Tien A.Y., Peng L., Lee S., Pearlson G.D. Structural differences in the cerebral cortex of healthy female and male subjects: a magnetic resonance imaging study // Psychiat. Res. - 1995. - Vol.61. - N3. - P.129-135.
Shaywitz B.A., Shaywitz S.E., Pugh K.R., Constable R.T., Skudlarski P., Fulbright R.K., Bronen R.A., Fletcher J.M., Shankweiler D.P., Katz L. et al. Sex differences in the functional organization of the brain for language // Nature. - 1995. - Vol.373. - N6515. -P.607-609
Silberman E., Weingartner H. Hemispheric lateralization of functions related to emotions // Brain Cognit. - 1986. - Vol.5. - P.322-353.
Sinton C.M. Sex differences in the interaction of emotional factors with memory // Intern. J. Neurosci. - 1981. - Vol.14. - P.211-218.
Skrandies W., Reik P., Kunze C. Topography of evoked brain activity during mental arithmetic and language tasks: sex differences // Neuropsychol. - 1999. - Vol.37. - N4. - P.421-430.
Smith M.L., Milner B., Residual memory deficits after unilateral cerebral excision // Annual Meet. Internat. Neuropsychol. Sympos. - Beaune, France, 1984. - P.10.
Snow W.G., Sheese S., Lateralizated brain damage, intelligence and memory: a failure to find sex differences // J. Consult. Clin. Psychol. - 1985. - Vol.53. - P.940-941.
Squire L.R., Alvarez P. Retrograde amnesia and memory consolidation: a neurobiological perspective// Current Opinion in Neurobiology. 1995. V.5.N2. P.169)
Steinmetz H., Jancke L., Kleinschmidt A., Schlaug G., Volkmann J., Huang Y. Sex but no hand difference in the isthmus of the corpus collosum // Neurology.- 1992. - Vol.42. - N4. - P.749-752.
Sternberg S. High-speed scanning in human memory // Science. - 1966. - Vol.153. - P.652-654.
Sternberg S. Memory scanning: mental processes revealed by reaction time experiments // Am. Sci. - 1969. - Vol.57. - P.421-457.
Sternberg S. Two operations in character recognition: some evidence from RT measurement // Percept. Psychophys. - 1976. - Vol.2. - P.421-457.
Taylor M.J., Smith M.L., Iron K.S. Event-related potential evidence of sex differences in verbal and nonverbal memory tasks // Neuropsychologia. - 1990. - Vol. 28. - N.7 - P.691-705.
Temple C.M., Cornish K.M. Recognition memory for words and faces in schoolchildren: a famale advantage for words //Brit. J. Develop. Psychol. - 1993. - Vol.11. - P.421-426.
Trenerry M.R., Jack C.R. Jr., Cascino G.D., Sharbrough F.W., Ivnik R.J. Gender differences in post-temporal lobectomy verbal memory and relationships between MRI hippocampal volumes and preoperative verbal memory // Epilepsy Res. - 1995. - Vol.20. - N1. - P69-76.
Tulving E., Kapur S., Markowitsch H.J., Craik F.I.M., Habib R., Houle S. Neuroanatomical correlates of retrievai in episodic memory: auditory sentence recognition // Proc. Natl. Acad. Sci. USA. 1994. - Vol.91. - N6. - P.2012-2016.
Vikingstad E.M., George K.P., Jonson A.F., Cao Y. Cortical language lateralization in right handed normal subjects using functional magnetic resonance imaging // J.Neurolog.Sci .- 2000. - Vol.17 . - P.17-27.
Wapner W., Hamby S., Gardner H. The role of the right hemisphere in the apprehension of complex linguistic materials // Brain Lang.- 1981.- Vol.14.-N1.- P.15-33.
Warrenburg S. Pagano R. Sex differences in EEG asymmetry predict performance in visiospatial tasks // Psychophysiol. - 1981. - Vol.18. - N2. - P.171-172.
Weinberger D.R., Gender differences in the normal lateralization of the supratemporal cortex - MRI surface -rendering morphometry of Heschl's // Cereb. Cortex . - 1994. - Vol.4 - N2. - P. 107 - 108.
Weiss S., Rappelsberger P. Long-range EEG synchronization during word encoding correlates with successful memory performance// Cognitive Brain Res. - 2000. - V9. - N3. - P.299-237.
Welsh T., Elliott D. Gender differences in a dichotic listening and movement task: lateralization or strategy? //Neuropsychol., 2001. – Vol . 39. - P. 25-35.
Wey T.V., Cook N.D., Landis T., Regard M. Lateralized lexical decisions and the effects of hemifield masks: a study of unterhemispheric inhibition and release // Intern. J. Neurosci. - 1993. - Vol.71. - P.37-44.
Wexler B.E., Lipman E. Sex differences in change over time in perceptual asymmetry // Neuropsychol. - 1988. - Vol.26 - P.943-948.
Wisniewski A. B. Sexually dimorphic partterns of cortical asymmetry, and the role for sex steroid hormones in determining cortical patterns of lateralization Psychoneuroendocrinol. – 1998. - Vol.23. - Is. 5. - P. 519-547.
Witelson S. F. Hand and sex diferences in the isthmus and genu of the human corpus callosum // Brain. - 1989. - Vol. 112. - P. 799-835.
Witelson S.F., Glezer I.I., Kigar D.L. Women have greater density of neurons in posterior temporal cortex // J. Neurosci. - 1995. - Vol.15. - P.3418-3428.
Witelson S. F. and Kigar D. L. Sylvian fissure morphology and asymmetry in men and women: Bilateral differences in relation to handedness in men. // The J. Compar. Neurol. – 1992. – Vol. 323. – P. 326-340.
Wittling W. Psychophysiological correlates of human brain asymmetry: blood pressure changes during lateralized presentation of an emotionally laden film // Neurophysiol. - 1990. - Vol.28. - P.457-470.
Wood F.B., Flowers D.L., Naylor C.E. Cerebral laterality in functional neuroimaging // Cerebral Laterality: Theory and Research. The Toledo Symposium. - Hillside, NJ: Erlbaum, - 1991. - P.103-115.
Zaidel E., Glarke, J.M., Suyenobu B. Hemispheric independence: a paradigm case for cognitive neuroscience // Neurobiology of Higher Cognitive Functions. - N.Y., 1990. - P.297-355.