АСИММЕТРИЯ НАПРАВЛЕНИЯ ДВИЖЕНИЯ У БЕСПОЗВОНОЧНЫХ
Г.П. Удалова, А.Я. Карась
Санкт-Петербургский Государственный Университет,
Московский Государственный Университет
По мере того как благодаря исследованиям ряда ученых (F. Nottebohm, 1979; V. Denenberg, 1983; В.Л. Бианки, 1985 и др.) становилось очевидным, что функциональная межполушарная асимметрия свойственная не только человеку, но также и высшим позвоночным, все более остро вставал вопрос о причинах возникновения этого свойства живых организмов и его развитии в эволюции. В этом аспекте логичным стало обращение внимания исследователей к проблеме латерализации функций у беспозвоночных. В сборнике “ Evolution and lateralization” П. Витт (Р. Witt, 1977) кажется впервые поставил эту проблему. Он, однако, не получил положительных данных о латерализации функций у пауков, изучая плетение паутины. Известно, что у позвоночных наиболее отчетливыми проявлениями функциональной межполушарной асимметрии являются моторные: предпочтение одной из конечностей, односторонние вращения , а также выбор определенного ( левого или правого ) направления движения. Все эти формы движения в той или иной степени связаны с ориентацией животного в пространстве. Выбор определенного направления движения как одной из форм ориентационного поведения, довольно хорошо изучен в связи с проблемой асимметрии функций ЦНС. Наличие такого рода пространственно-моторной асимметрии показано у нескольких видов позвоночных: крыс (S. Glick, R. Cox, 1976; S. Glick et al. 1977; J. Hoffman, 1980; S. Glick, D. Ross, 1981; D. Ross, S. Glick, 1981; E.A. Рябинская, 1982; E.A. Рябинская, T.C. Валуйская, 1983 и др.), мышей (B.B. Тхоржевский, 1973; A. Korczyn, Y. Eshel, 1979; Г.П. Удалова, 1996; Н.В. Маркина и др., 1999), хомяков (Т. Giehrl, Н. Distel, 1980), кошек (S. Glick, L. Weaner, R. Meibach, 1981; S. Glick, D. Ross, 1982; A.A. Дутов и др., 1985), а также у рыб (C.И. Глейзер, 1981; В.А. Непомнящих, В.А. Гремячих, 1993). В экспериментах на грызунах выявлено неодинаковое значение корковых и подкорковых отделов правого и левого полушарий головного мозга для формирования одностороннего предпочтения в различных экспериментальных условиях: при обучении в лабиринте, при альтернативном выборе зрительных стимулов, в тесте “открытое поле“ (G. Sherman et al., 1980; В.Л. Бианки и др., 1982; Г.П. Удалова, В.В. Михеев, 1982; Г.П. Удалова, 1983, 1988; Т. Moran et al., 1984; К. Kubos, R. Robinsоn, 1984; Г.П. Удалова, И.А. Кашина, 1987). Изучались механизмы возникновения у беспозвоночных асимметрии направления движения, в том числе и нейрохимические (S. Glick et al., 1977; A.A. Дутов, C.C. Анохов, 1983; Л.Л. Клименко и др., 1995).
Что же касается беспозвоночных, то ещё сравнительно недавно факты, свидетельствующие о пространственно-моторной асимметрии у них можно было найти в исследованиях, где проблема функциональной асимметрии ЦНС по-существу не ставилась. Находки такого рода носили вторичный характер как дополнение к какой-либо основной теме. Из известных нам материалов упомянем работу, в которой было показано передвижение летучей саранчи преимущественно направо или налево, что рассматривалось как результат асимметрии моторного контроля (D. Wilson, 1968).Стойкий, независящий от внешних факторов выбор левого или правого направления полета при поисках пищи выявился у шмелей Bombus lapidarius (J. Cheverton, 1982).Эти данные указывали на наличие индивидуального одностороннего предпочтения (левого или правого), но вопрос о возможности выявления у беспозвоночных видоспецифической функциональной асимметрии оставался открытым. Систематическое, целенаправленное исследование пространственно-моторной асимметрии у беспозвоночных было начато в лаборатории В.Л. Бианки (1985) как одно из важных направлений разработки проблемы функциональной латерализации ЦНС у животных. В настоящее время имеются условия для обобщения экспериментального материала, полученного при изучении предпочтения направления движения у беспозвоночных, начиная с низших (черви) и кончая высшими общественными насекомыми (осы, пчелы, муравьи).
Наиболее примитивными животными с билатеральным строением тела и нервной системы являются плоские ресничные черви. Возникновение билатеральной симметрии у этих животных произошло в процессе эволюции при переходе от плавания к передвижению по твердому субстрату (Л.Н. Cеравин, А.А. Карпенко, 1987). Ресничные черви дали начало всему многообразию зеркально-симметричных организмов (А.В. Иванов, Ю.В. Мамкаев, 1973). Особенности строения турбеллярий отразились на характере восприятия ими окружающего пространства (Л.Н. Серавин, А.А. Карпенко, 1987). Показано, что турбеллярии (Сonvoluta convoluta) отчетливо различают (в отличие от инфузорий) стимулы, действующие на них справа или слева. Повороты переднего конца тела влево или вправо после сильных раздражений не являются для отдельной особи четко равновероятными, что, по мнению авторов исследования, согласуется с гипотезой об индивидуальном предпочтении левого или правого направления движения у планарии Dendrcoelum lacteum (G. Chapounthier et al., 1968). Специально изучали асимметрию направления движения у ресничных червей, наблюдая за ориентацией в Т-образном лабиринте пресноводных планарий четырех видов: Dugestia tigrina (бесполая и половая расы), Dugestia lugubris, Ijimia tennuis, Blellacephala punctata (В.Л. Бианки, И.М. Шейман, Е.В. Зубина, 1990). Планарию помещали в стартовый коридор головным концом в направлении развилки лабиринта. После свободного выбора ею одного из направления червя вновь переносили в стартовый коридор. Серию из 10 проб прерывали на час, а затем серии повторяли до 50 проб. Для элиминации сигнального значения выделяемой планариями слизи лабиринт протирали спиртом и водой. При анализе индивидуального поведения планарий выявили особей без достоверного выбора одного из направлений (“симметрики”) и с преимущественным выбором правого или левого направления.”Симметрики “составили около 60% выборки. Среди “асимметриков“ число “правшей“ и “левшей “ было неодинаковым у разных видов планарий. У Dugestia tigrina “правши “ составили 23-32% , а “левши“- 10%. У трех других видов “левшей“оказалось больше, чем “правшей“: соответственно 22-40%; и 9-11%. По суммарному количеству односторонних поворотов в целом для популяции выявилось достоверное предпочтение правых выборов у Dugestia tigrina ( соответственно 56% правых поворотов и 44% левых, р<0,05). У остальных трех видов наблюдалась лишь тенденция к преобладанию правых или левых выборов. На бесполой расе Dugestia tigrina изучали корреляцию между первым и последующими поворотами. Суммарные количества первых правых или левых поворотов различались незначительно, хотя и с тенденцией в сторону преобладания правых поворотов, которая была бодее выраженной у “асимметриков”, чем у” симметриков”. Примерно у 2/3 особей “правшей” первая реакция была правосторонней и у 1/3 – левосторонней, а у “левшей “ соотношение было таким же, но с обратным знаком направления. Обнаруженная у планарий асимметрия характеризовалась определенной устойчивостью, что проявилось , в частности, в отсутствии ее сдвигов в звисимости от сезона, а также от ориентации лабиринта по сторонам света.
Итак, у представителей низших многоклеточных беспозвоночных – планарий четырех видов “симметричные” особи преобладают над” асимметричными”, а последние подразделяются на группы с преобладанием правых или левых поворотов. Такого рода предпочтение направления движения было обозначено как примитивная форма функциональной видовой асимметрии, а преобладание левого или правого выбора для асимметричной части популяции у планарий рассматривается в качестве видового признака. Существенен также вывод о том, что функциональная асимметрия возникает у червей одновременно с морфологической билатеральной симметрией. Доказательством возникновения функциональной асимметрии на столь раннем уровне филогенеза беспозвоночных служат и данные о преобладании особей с левосторонними вращательними движениями у морских турбеллярий трех видов Pseudostomun quadrioculatum, Coronopharyns pusillus и Provortex karlingi (К.К. Богута, 1985). Вместе с тем, К.К. Богута показал, что этим турбелляриям не свойственна точная зеркальная симметрия нервной системы.
До сих единичны работы по исследованию пространственно-моторной асимметрии у моллюсков и ракообразных. Правосторонняя асимметрия обнаружена у брюхоногого моллюска Hyanassa obsoleta при тестировании в T-образном лабиринте (D. Bretz, R. Dimock, 1983).
У ракообразных е уровень симметризации нервной системы выше, чем у ресничных червей. Например, речного рака уже строго симметрично строение нейропиля грудных ганглиев. У речных раков Astacus leptodactylus обнаружено достоверное правостороннее предпочтение при ориентации в восьмилучевом лабиринте и на специальных полигонах (Г.В. Баранюк и др., 1983).
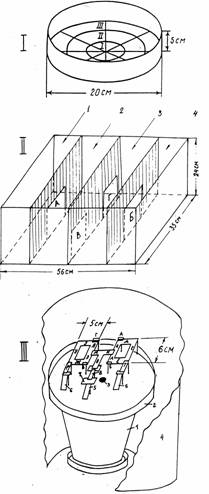
Нами в качестве объектов для изучения асимметрии направления движения использовались представители ракообразных с разной степенью сложности строения нервной системы и поведения: бокоплавы (Gammarus oceanicus ) и черноморские крабы ( Carcinus meanas ). Эксперименты на гаммарусах Белого моря проводили по методике “открытого поля” (Г.П. Удалова, А.Я. Карась, М.И. Жуковская, 1990 а, б). Аквариум (рис. 1,1) наполняли 800 мл морской воды до уровня 4,5 см.
Рис. 1. Схемы экспериментальных установок: 1 – для тестирования гаммарусов;
11 – для изучения реакции избегания у крабов; III – для выработки лабиринтного навыка у муравьев. На рис. 1. III: 1 – гнездо, 2 – арена, 3 – вход в гнездо, 4 – ширма, 5 – центральный мостик, 6 - боковые мостики, 7 – стартовая площадка, 8 – съемный мостик; О – вход в лабиринт, А и Г - целевые площадки, Б и В – симметричные пустые площадки, Т и Р – выходы; ИКДСЕЛ и МЯЭУЖЗ – остальные участки правой и левой половин лабиринта.
Его дно было разделено на три зоны и на 8 секторов (рис.1,1). Тестировали наиболее активных, крупных особей старшего возраста размером 2- 2,5 см. Перед проведением опыта каждого гаммаруса переносили из большой ванны в сосуд объемом 200 мл. В начале тестирования животное помещали в центр аквариума и после одноминутной адаптации наблюдали его двигательную активность в течение шести тридцатисекундных интервалов. Оценивали особенности плавания, учитывая направление движения, нахождение в каждой из трех зон, переворачивания и остановки. Каждый переход из одного сектора в соседний или из одной зоны в близлежащую принимали за одно движение. Проводили 1-3 опыта с часовыми интервалами, во время которых бокоплава отсаживали в большой сосуд. Эксперименты осуществляли днем при естественном освещении. Асимметрию выявляли, сопоставляя число движений в зоне III в направлении по часовой стрелке и против нее. Рассчитывали коэффициент асимметрии (Кас) как отношение разности этих показателей к их сумме. Определяли число особей, двигавшихся преимущественно по часовой стрелке или против нее (обозначим их ”правшами” и ”левшами”) или без достоверного различия между этими величинами (“симметрики”). Статистическую обработку данных здесь, как и в последующих разделах наших исследований, производили используя стандартные методы Стьюдента- Фишера, а также критерии: парный, хи-квадрат и Вилкоксона.
В целом для всех исследованных животных (204 бокоплава) обнаружилась пространственно-моторная асимметрия (таблица 1 ).
|
№ серии |
Число особей |
I |
II |
II |
Остановки |
III направо |
III налево |
Кас |
|
1 |
60 |
14 |
48 |
8041 |
210 |
5643 |
2396 |
0,404 |
|
2 |
76 |
40 |
160 |
9535 |
65 |
6158 |
3377 |
0,292 |
|
3 |
68 |
55 |
155 |
8811 |
153 |
3973 |
4838 |
-0,098 |
|
Сумма |
204 |
109 |
363 |
26387 |
428 |
15776 |
10611 |
0,196 |
Таблица 1. Количество движений и остановок, а такжеи пространственно-моторной асимметрии у бокоплавов в первом опыте каждой серии и, в целом, для всех животных. I, II, III – зоны.
Она проявилась в достоверно большем количестве движений по часовой стрелке (направо), чем против нее (налево). Животные с правосторонней или левосторонней асимметрией преобладали, составив 93,5% выборки. Достоверным (р<0,05) оказалось также большее число особей с правосторонней асимметрией. Двигательная активность и асимметрия изменялись при повторных тестированиях. В целом для исследованной группы из 38 особей в первом тесте наблюдалось правостороннее предпочтение ( Кас= + 0,055; р 0,001) , во втором - левостороннее ( Кас= -0,110; р 0,001), а в третьем тесте вновь появилась правосторонняя асимметрия, причем более сильная ( Кас=+0,208; р 0,001). В ходе эксперимента возрастала абсолютная величина Кас. При первом тестировании было 20 ”правшей’’, 17’’левшей’’и один ’’симметрик”, при втором- соответственно 16, 20 и 2 особи, при третьем- 21, 14 и 3. ( рис. 2, 1, 11 ). Изменения Кас, а также двигательной активности, у ”правшей” и “левшей” проявлялись в неодинаковой степени. У ”правшей” двигательная активность усиливалась во втором опыте и снижалась в третьем. Асимметрия сначала резко ослабевала, а затем усиливалась, не достигая однако исходного уровня (рис.2,1, А ).
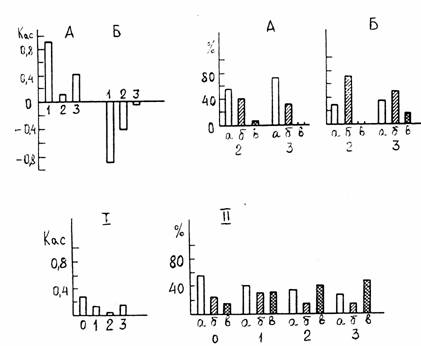
Рис. 2. Показатели пространственно-моторной асимметрии у гаммарусов.
Сверху: асимметрия у особей с исходным предпочтением движения по часовой стрелке (А) и против нее (Б) при повторных тестированиях: 1 – коэффициент асимметрии, 11 – число “правшей” (а), “левшей” (б) и “амбидекстров”(в) в выборке (%) 1, 2, 3 – первый, второй, третий опыты.
Внизу: изменение асимметрии при действии токсиканта.
1 – коэффициент асимметрии у всей выборки ( n =40 );
11 – число “правшей” (а), “левшей” (б) и “амбидекстров” (в) в выборке (%). 0 – опыт до применения нефтяного загрязнения, 1, 2, 3 – опыты после интоксикации.
У “левшей” повторные тестирования приводили к постепенному ослаблению двигательной активности и асимметрии (рис. 2, 1, Б ). Среди” правшей” и среди “левшей” при повторных тестированиях появились особи с инверсией исходного знака асимметрии и “ симметрики” (рис.2,11 ). Итак, асимметрия у бокоплавов характеризовалась динамичностью, причем “правши” оказались более устойчивыми к повторным тестам, чем “левши”. Динамичность асимметрии особенно отчетливо проявилась, когда тесты проводили 2-3 раза на фоне нефтяного загрязнения. При действии токсиканта резко снижалась двигательная активность. Исходная положительная величина коэффициента асимметрии уменьшалась ( рис. 2, 11). В ходе эксперимента “асимметриков”, особенно “правшей” стало меньше, а “симметриков” - больше.” Правши” оказались менее устойчивыми к действию токсиканта,чем “левши”, если судить по. изменению величины Кас, но более устойчивыми по другому показателю- сохранению у каждой особи исходного знака асимметрии (он не менялся у 30% среди” правшей”, а среди” левшей” только у 14%).
У крабов асимметрию направления движения изучали на 54 особях – самцах с диаметром карапакса 6,2- 7,7 см по методике активного избегания (А.Я. Карась, Г.П. Удалова, 1988). Экспериментальная установка состояла из аквариума, разделенного перегородками на четыре отсека (рис.1, 11).Стартовый отсек 1 и отсек 2, отсеки 3 и 4 соединялись центральными проемами (соответственно А и Б ) симметричными продольной оси аквариума. Из отсека 2 (зона выбора) в отсек 3 вели боковые проемы: правый В и левый Г. Отсеки 3 и 4 затемнялись и служили зонами комфорта для крабов этого вида, обладающих отрицательным фототаксисом. Условия опыта обеспечивали равновероятный выбор правого или левого направления. Опыты проводили при диффузном естественном освещении в июне при температуре 22-24 градуса. Каждое животное тестировали в течение одного дня с 9 до 19 часов.После того как краба помещали в отсек 1 напротив проема А, он выходил в отсек 2 и через проем В или Г попадал в отсек 3 и далее через проем Б в отсек 4 ( убежище). Краб не менее двух минут оставался в отсеке 4, а затем его снова помещали в отсек 1. На 21 особи ставили один опыт ( по 10 проб), . на 33 крабах – по два опыта ( каждый по 10 проб), разделенных часовым интервалом. О наличии асимметрии судили по соотношению числа выходов через проемы В и Г, а также по количеству особей с относительно большим числом правых и левых выборов.
Отчетливое правостороннее предпочтение, т.е. большее число выходов в отсек 3 через правый проем В, а не левый Г, выявилось в целом для обоих выборок ( таблица 2).
|
Группы |
a |
б |
Уровень значимости |
|
I (n=21) |
6,09+0,4 |
3,9+0,4 |
<0,01 |
|
II (n=33) |
12,5+0,4 |
7,2+0,5 |
<0,001 |
|
III (n=54) |
6,3+0,2 |
3,7+0,2 |
<0,001 |
|
«правши» (n=33) |
9,1+0,2 |
3,4+0,2 |
<0,001 |
|
«левши» (n=12) |
3,3+0,2 |
6,8+0,2 |
<0,001 |
Таблица 2. Среднее число правых ( а) и левых ( б ) выборов у группы крабов с одним опытом ( 1 ), с двумя опытами (11) и у всех особей в целом (111) Приведены суммарные и средние для каждой группы значения; п = число животных.
Коэффициент асимметрии для правшей - 0,46, для «левшей» - -0,35.
Первая выборка состояла из 12”правшей” , 3” левшей “и 6 “симметриков”; вторая - из 24, 5 и 4 особей соответственно. В целом” правшей” оказалось в 4,5 раза больше, чем “левшей”( р 0,01). Среди 54 крабов было выявлено 22 особи с достоверным различием числа правых и левых выборов, причем среди асимметриков 20 крабов имели правостороннее предпочтение и только 2 – левостороннее. Правосторонняя асимметрия в целом для всех крабов обнаружилась уже во второй пробе. В дальнейшем она сохранялась с достоверностью или как тенденция. Правосторонняя асимметрия в сумме для первого десятка проб была несколько сильнее, чем для второго. Различие у крабов размеров клешней позволило рассмотреть вопрос о взаимосвязи морфологической асимметрии и функциональной асимметрии направления движения. У 43 крабов большей была правая клешня, а у 11 – левая. В каждой группе отмечалась правосторонняя асимметрия, причем во второй – несколько более сильная: соотношение правых и левых выборов равнялось соответственно 1,6 и 2,7, т.е. особи с большей левой клешней относительно чаще выбирали правый проем, чем крабы с большей правой клешней. Обнаружилась определенная зависимость правого или левого выбора от начальной позы животного в стартовом отсеке 1. Когда краб ориентировался правой стороной тела и правой клешней ( безразлично большой или малой) к правому проему (B), правые выборы наблюдались в 88% случаев в первом десятке проб и в 84% - во втором десятке. Если крабы были первоначально сориентированы левой стороной к левому проему (С), левый проем выбирался соответственно в 86 % и 81% .
Насекомые представляют еще один класс беспозвоночных, характеризуемый сложным симметричным строением нервной системы и высокоорганизованным поведением. Выше уже упоминались факты, указывающие на возможность выявления моторной асимметрии у насекомых. Впервые, наверное, С.D. Putnam (1962), изучая поведение жуков Aleochara bilineata в Y- образном лабиринте, обнаружил в исследованной выборке более половины особей, предпочитавших достоверно выбирать левое или правое направление. В целом для выборки, однако, асимметрия определенного знака не была показана. В этой работе была прослежена зависимость каждого последующего выбора от нескольких предшествовавших, в том числе и от первого в опыте выбора. Рассматривалась даже, хотя и безрезультатно, возможность выявления половых различий в отношение предпочтения право- или левостроннего направления движения в лабиринте.
В.Л. Бианки и И.М. Шейман (1985) изучали в Т-образном лабиринте поворотные тенденции у мучного хрущака (Tenebrio molitor) причем как на личинках, так и на имаго. Затемнение и отруби в обоих коленах лабиринта служили подкреплением. Для выявления асимметрии использовали три критерия: направленность первого поворота; обучение в лабиринте в течение 100 проб у жуков и 50-для личинок; переделка первоначально предпочитаемого направления движения в лабиринте. В последнем случае за неправильный выбор животное наказывалось механическим ударом. По первому повороту в выборке из 130 личинок 53% особей поворачивали направо, а 47% - налево, т.е. достоверного предпочтения направления движения не выявилось. В другой выборке из 60 личинок у 60% особей наблюдался первый правый поворот. В целом, можно было говорить о тенденции к правшеству. По результатам экспериментов с обучением и переделками выбор правых и левых поворотов оказался равновероятным как у личинок ( 40 особей), так и у жуков ( 20 особей). Было обнаружено, однако, что прочность левосторониих исходных поворотных тенденций более выражена по сравнению с правосторонними. Этот феномен проявлялся как в условиях свободного выбора (у личинок и жуков), так и при переделке исходного направления движения на противоположное (у личинок). Авторы работы пришли к выводу о том, что такого рода различия указывают на существование у мучного хрущака некоторых форм функциональной асимметрии. Здесь следует упомянуть о том, что у мучного хрущака индивидуальная функциональная асимметрия может иметь морфологическую основу. Показано (F. Brihschneider, 1914), что у личинок и взрослых особей этого вида грибовидные тела асимметричны: у 2/3 насекомых левая доля заходит за правую, а у 1/3 –правая доля заходит за левую.
Исследовалась асимметрия направления движения и на более высокоорганизованных насекомых: осах, пчелах и муравьях. Было замечено, что пчелы и осы, обследуя группу близко расположенных объектов, и многократно возвращаясь за приманкой, запоминают наиболее простой правильный путь облета, чаще всего по кругу, причем двигаясь преимущественно против часовой стрелки, т.е. налево (Г.А. Мазохин-Поршняков, В.М. Карцев, 1979). Поворот пчел преимущественно налево наблюдался и в другом исследовании (Р. Schmid-Hempel, 1984). Позже в специально проведенных экспериментах изучали (В.М. Карцев и др., 1988) асимметрию траекторий движения, выбираемых медоносными пчелами (Apis mellifora) и складчатокрылыми осами ( Paravespula spp.) при поиске корма. В опытах во время фуражировочного облета насекомому надо было по одному разу посетить каждую из четырех кормушек с сахарным сиропом. Анализировали только случаи, когда пчела или оса, прилетевшая за сиропом, собирала корм из четырех кормушек, не возвращаясь на посещавшиеся ранее. Исключали возможность ориентации насекомых по каким-либо удаленным наземным вехам. Подтвердилось достоверное преобладание в выборках особей, предпочитавших летать против часовой стрелки. При этом, обсуждая этологический механизм асимметрии траекторий движения, отметили следующее: если представить себе насекомое, по прямой подлетающее к середине столика с четырьмя кормушками по углам, то чтобы двигаться против часовой стрелки, т.е. поворачивать налево, ему надо сначала повернуть направо. По результатам обследования значительного количества пчел и ос были сделаны следующие заключения:
1) по крайней мере рабочим особям общественных насекомых присуща моторная асимметрия; 2) видовую моторную асимметрию можно расценить как свидетельство функциональной асимметрии головного мозга, тогда как индивидуальную (групповую) асимметрию вполне можно объяснить лишь повторением насекомым своего пути, первый раз найденного случайно.
Перейдем теперь к изложению результатов исследования пространственно-моторной асимметрии у муравьев.
Семью муравьев (Myrmica rubra), состоявшую из рабочих особей, самок и расплода, помещали в искусственное гнездо за 3-4 недели до начала опытов. ( рис.1, III ). Над гнездом находилась арена (2), на которой устанавливали лабиринт из съемных бумажных мостиков. Он состоял из двух симметричных половин, каждая из которых включала целевую площадку ( А или Г) с подкреплением , симметричную ей пустую площадку ( Б или В ) и выход (Т или Р ). Лабиринт соединялся с ареной центральным мостиком (5) и двумя боковыми (6). Подкрепление: куколки собственного вида или капля сахарного сиропа находидось на обеих целевых площадках. Муравьев-разведчиков сначала приучали подниматься на стартовую площадку (7), помещая на ней подкрепление. После этого у наиболее активных особей, помеченных индивидуально, вырабатывали лабиринтный навык транспортировки расплода или фуражировки. В первом случае реализовалась социальная защитная мотивация, во втором -социальная пищевая. Как только один из муравьев проходил со стартовой площадки на первый участок лабиринта (О), съемный мостик (8) удаляли. Передвигаясь по лабиринту, муравей мог взять подкрепление, а затем через один из выходов выйти на арену и далее в гнездо. Траекторию передвижения от входа в лабиринт до выхода из него (цикл) записывали последовательностью букв, обозначавших участки лабиринта. Прохождение каждого участка принимали за единицу движения. После каждого цикла все части лабиринта заменяли. Траектории передвижения и продолжительность нахождения в лабиринте, а также в значительной степени частота заходов в него определялись самими муравьями. Конструкция лабиринта позволяла муравьям реализовывать множество правильных траекторий : минимальной длины или избыточных, т.е. с лишними и /или повторными подходами к участкам лабиринта. Правильными циклами ( решениями ) считали такие, в которых муравей впервые брал подкрепление, не заходя предварительно на площадку Б и/или В. Эксперимент на каждом муравье продолжался 1-3 дня. Формирование лабиринтного навыка оценивали по совокупности показателей: возникновению первого правильного решения, количеству правильных решений, стабилизации навыка, среднему числу движений в цикле и на его первом этапе (от входа в лабиринт до первого взятия подкрепления), отношению числа подходов к целевым площадкам со взятием подкрепления и исследовательских подходов к ним, а также по другим критериям. Асимметрию определяли по числу подходов к целевым площадкам с подкреплением, подходам к площадкам Б и В, выходам из лабиринта и суммарному числу движений на всех участках правой или левой половин лабиринта и по другим показателям.. У каждой особи и в целом для выборок рассчитывали коэффициент асимметрии как частное от деления разности соответствующих показателей для правой и левой половин лабиринта к их сумме.
Стратегии поведения муравьев при обучении в многоальтернативном лабиринте зависели от вида мотивации (забота о потомстве или пищевая) и уровня пищевой мотивации (А.Я. Карась, Г.П. Удалова, Е.В. Загораева, 1986; А.Я. Карась, Г.П. Удалова, Б.А. Дашевский, 1986; А.Я. Карась, Г.П. Удалова, 1998, 2000; Г.П. Удалова, А.Я. Карась, 1999 и др.). При мотивации заботы о потомстве, а также при высоком уровне пищевой мотивации выявился эффект сверхбыстрого обучения. Стохастическое, неоптимизированное поведение в лабиринте наблюдалось при низкой пищевой мотивации. Однако во всех экспериментальных моделях выявлялась пространственно-моторная асимметрия либо в индивидуальной форме, либо в целом для выборок (Г.П. Удалова, А.Я. Карась, 1985, 1986, 1989 а, б; А.Я. Карась, Г.П. Удалова, Е.В. Загораева, 1986; А.Я. Карась, Г.П. Удалова, Б.А. Дашевский 1986 и др.). Асимметрию, обнаруженную для достаточно больших выборок, обозначали как видовую или “ семейную “. При защитной мотивации асимметрия оказалась более выраженной, чем при пищевой, особенно при сниженной потребности семьи в пище. На таблице 3 представлены суммарные данные для 140 муравьев из 7 семей, переносивших расплод и для 30 муравьев из 3 семей, обучавшихся фуражировке при относительно низком уровне пищевой мотивации. В первом случае достоверная правосторонняя асимметрия проявилась по подходам к целевым площадкам, по выходам, по числу “ чистых” циклов ( т.е. траекторий, проходивших только на одной половине лабиринта ), а также по суммарному числу движений на каждой половине лабиринта. Во втором случае почти все показатели асимметрии были хуже, причем достоверное правостороннее предпочтение обнаружилось по подходам к площадкам Б и В, по “смешанным” циклам и суммарному числу движений, но отсутствовало по таким важным показателям, как подходы к целевым площадкам и выходы. У всех животных в целом выявилось правостороннее предпочтение по пяти из семи показателям. Достоверная индивидуальная асимметрия отмечалась у 57% особей при защитной мотивации и только у 26% при пищевой, а по суммарному числу движений соответственно у 84% и 60%. При выработке навыка фуражировки на фоне высокой пищевой мотивации семьи асимметрия проявлялась отчетливее, но ее показатели в целом были хуже, чем при защитной мотивации (А.Я. Карась, Г.П. Удалова, 1998, 2000).
Муравьи разных семей различались по знаку и степени выраженности асимметрии , а также по динамике ее показателей в ходе обучения (Г.П. Удалова, А.Я. Карась, 1989 а, б). На рис.3 представлена динамика суммарного числа движений на правой (1) и левой (2) половинах лабиринта у муравьев из трех разных семей, обучавшихся при низкой пищевой мотивации переносить расплод (а, б) или фуражировке (в).
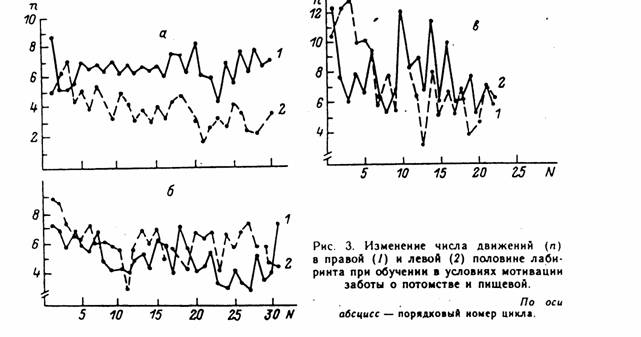
Рис. 3. Динамика числа движений на правой (1) и левой (2) половинах лабиринта при обучении муравьев транспортировке расплода (а, б) или фуражировке (в).
По абсциссе – порядковый номер цикла, по ординате – правосторонние (1) и левосторонние (2) движения для соответствующего цикла. На каждом графике указаны усредненные для всего периода обучения значения правых (1) и левых (2) движений.
В первом случае (а) правосторонняя асимметрия возникла уже в четвертом цикле, затем сохранялась и усиливалась. Аналогичным образом проявилась динамика в отношении подходов к целевым площадкам и выходам. Во втором случае (б) была левосторонняя асимметрия, причем более вариабельная, чем в первом случае. Знак и степень выраженности показателей асимметрии менялись весьма значительно при обучении фуражировке (в).
Асимметрия направления движения оказалась в целом более отчетливой и стабильной у муравьев-“правшей”, чем у” левшей”. Так, на рис.4 приведены суммарные данные для 6 муравьев-“правшей” и 5 – “левшей” из разных семей, обучавшихся переносу расплода в течение 60 циклов.
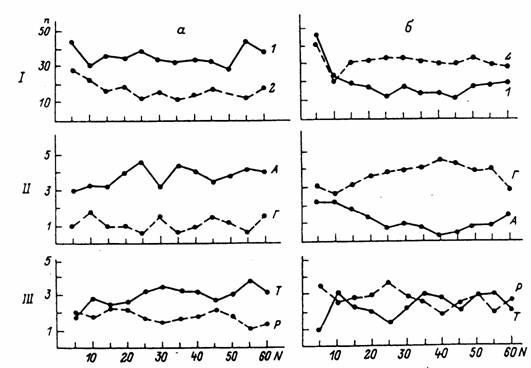
Рис. 4. Динамика пространствено-моторной асимметрии при обучении
Муравьев транспортировки расплода у группы особей с правосторонним (а) и левосторонним (б) предпочтением.
Показатели асимметрии: 1 - суммарное количество движений на правой (1) или левой (2) половине лабиринта, 11 - число подходов к целевым площадкам А или Г со взятием подкрепления, III – число выходов Т или Р.
У ”правшей” асимметрия оказалась весьма стабильной по всем трем показателям (рис.4, а ). У ”левшей” она была менее выраженной, а по числу выходов не выявилась (рис.4,б ). Подчеркнем, что окончательное закрепление знака асимметрии обычно совпадало со стабилизацией навыка.
Показатели асимметрии, как правило, изменялись в экзаменах, в которых менялись тип мотивации, уровень пищевой мотивации или структура лабиринта. Формирование новой стратегии поведения в этих условиях сопровождалось увеличением, снижением или инверсией асимметрии в соответствии со спецификой изменившейся экспериментальной ситуации. Следовательно, пластичность поведения муравьев может проявляться не только в быстрой и адекватной модификации сформированного при обучении навыка, но также в изменении пространственно-моторной асимметрии. В качестве примера приведем результаты экспериментов со множественными переделками навыка транспортировки расплода (Г.П. Удалова, М.И. Жуковская, А.Я. Карась, 1991, 1992). При обучении подкрепление находилось на обеих целевых площадках в одной серии (21 муравей) или на одной из них ( 22 муравья). В конце обучения у каждого муравья выявилась индивидуальная право- или левосторонняя асимметрия. В экзамене подкрепление помещали поочередно на предпочитаемую или непредпочитаемую площадку. Муравьи оказались способными выполнять до 8 переделок ранее выработанного навыка. Сформированные при обучении минимизированные асимметричные траектории (“чистые циклы”) преобразовывались в противоположные по направлению, очевидно, по механизму зеркального переноса. Коренным образом перестраивалась часть траекторий, связанная непосредственно с выбором целевой площадки. Первоначальная асимметрия по числу подходов к целевым площадкам или” чистых” циклов изменялась адекватно новым условиям. По другим показателям асимметрия сохранялась по знаку или же не выявлялась с достоверностью. В целом асимметрия ослабевала. Изменение знака исходного предпочтения зависело от условий обучения. Последние переделки муравьи выполняли лучше, чем первые, причем особенно те особи, у которых при обучении сформировалось предпочтение левой, а не правой, целевой площадки.
Существенные или относительно небольшие изменения исходной пространственно-моторной асимметрии, образовавшейся при обучении транспортировки расплода, прослеживались и в других вариантах экзаменов со сменой местоположение подкрепления (Б.А. Дашевский, А.Я. Карась, Г.П. Удалова, 1989) или с поворотом лабиринта в горизонтальной плоскости (А.Я. Карась, Г.П. Удалова, Б.А. Дашевский, 1995) или в вертикальной (Г.П. Удалова, А.Я. Карась, 1997).
Приведем еще данные опытов, в которых муравьев обучали навыку фуражировки при низком уровне пищевой мотивации, а экзамен проводили на фоне пищевой депривации семьи (А.Я. Карась, Г.П. Удалова, 1998). При обучении достоверная асимметрия ( правосторонняя ) выявилась только по суммарному числу движений ( Кас= + 0,083). В экзамене асимметрия обнаружилась по всем трем основным показателям: подходам к целевым площадкам, выходам и суммарному числу движений ( Кас равнялся соответственно +0,476; + 0,186 и + 0,435). Такое усиление асимметрии сочетались с быстрой минимизации траекторий и стабилизацией лабиринтного навыка, что было адекватно и биологически оправдано.
Для проверки предположения о том, что одностороннее моторное предпочтение может закрепляться после первого, возможно случайного, одноименного поворота в симметричной среде, рассматривали в целом у всех муравьев взаимосвязь между первым правым ( ОИ ) или левым (ОМ ) поворотом в первом цикле и основными показателями асимметрии (Г.П. Удалова, А.Я. Карась, 1989 а). По этим поворотам обнаружилось отчетливое правостороннее предпочтение: правый поворот – выявился у 57% особей, а левый - у 43%. Исходное “правшество” чаще приводило в конце обучения к формированию правосторонней, а не левосторонней асимметрии по суммарному числу движений ( р<0,05 ) и по выходам из лабиринта (р 0,01), но не по подходам к целевым площадкам. При исходном ”левшестве” наблюдалось практически равновероятное закрепление в процесс обучения правого или левого направления движения. Полученные данные пока не дают основания говорить об особом значении первого выбора направления движения для окончательного формирования в ходе обучения знака асимметрии. В то же время они подчеркивают наличие правосторонних тенденций при пространственной ориентации муравьев.
Заключение
Обобщая имеющийся в настоящее время экспериментальный материал можно сделать вывод о возможности выявления у беспозвоночных индивидуальной, групповой (популяционной), а также видовой асимметрии направления движения. Асимметрия такого рода обнаружена у животных весьма далеких таксономических групп, причем при реализации простых или довольно сложных форм поведения: тигмотаксис у гаммарусов, реакция активного избегания у крабов, обучение в лабиринте у планарий, жуков, речных раков и муравьев, фуражировочный облет у ос, шмелей и пчел.
Если у планарий и мучного хрущака предпочтение направления движения проявляется в довольно примитивной форме функциональной видовой асимметрии (В.Л. Бианки, И.М. Шейман, 1985; В.Л. Бианки и др., 1990), а у пчел и ос она выражается в предпочтении облета кормушек против часовой стрелки, т.е. в левостороннем направлении (Г.А. Мазохин-Поршняков, В.М. Карцев, 1979; В.М. Карцев и др.,1988), то, по нашим данным, у гаммарусов, крабов и муравьев, а также у раков (Г.В. Баранюк и др., 1983) отмечается отчетливая видоспецифическая правосторонняя асимметрия.
Очевидна зависимость показателей обнаруженной асимметрии от специфических условий эксперимента, характера поведенческой реакции и числа проб и других факторов. У муравьев отчетливо проявилось значение типа мотивации и ее уровня, соотношения исследовательской и доминирующей мотиваций. Так, сильная исследовательская активность ослабляет асимметрию. Семейные, а также индивидуальные особенности асимметрии у муравьев могут зависеть от потребности семьи в пище, общедвигательной активности, а также условий среды обитания. Указывают, например, что одни муравьи функционально связаны с левой половиной гнезда, а другие особи - с правой (А.А. Захаров, Т.Н. Орлов, 1984). Факторами, мешающими выявить присущую, возможно, генетически обусловленную, асимметрию направления движения у животных, могут служить также недостаточно однородная среда, присутствие внешних ориентиров (В.А. Непомнящих, В.А. Гремячих, 1993) или наличие токсических веществ (Г.П. Удалова и др., 1995).
Очевидно, что для надежного выявления знака и степени выраженности асимметрии целесообразно применять совокупность поведенческих показателей что особенно ясно продемонстировали эксперименты на муравьях.
Тот факт, что у представителей различных таксономических групп беспозвоночных: гаммарусах, крабах, речных раках, моллюсках и муравьях, а также у позвоночных животных (G. Sherman et al., 1980; J. Hoffman, 1980; S. Glick, D. Ross, 1981; Г.П. Удалова, В.В. Михеев, 1982; Г.П. Удалова, И.А. Кашина, 1987; Г.П. Удалова, 1996 и др.), в сходных экспериментальных условиях выявилась правосторонняя асимметрия направления движения, свидетельствует о существовании каких-то общих филогенетически обусловленных механизмов, обеспечивающих возникновение пространственно-моторной асимметрии. Выявление у некоторых видов беспозвоночных : пчелы, осы, планарии (В.М. Карцев и др., 1988; В.Л. Бианки и др., 1990) левосторонней асимметрии оставляет открытым вопрос о соотношении генетических и средовых факторов в формировании пространственно-моторного предпочтения.
В опытах на крысах было показано, что предпочтение одного из направлений при обходе радиального лабиринта уменьшало число ошибочных посещений уже обследованных коридоров (Е.А. Рябинская, Т.С. Валуйская, 1983). Однако не во всех экспериментальных ситуациях можно однозначно определить даже у позвоночных функциональное значение асимметрии направления движения (Г.П. Удалова, В.В. Михеев, 1988; В.А. Непомнящих, В.А. Гремячих, 1993). Противоречивы и данные (J. Cheverton, 1982; В.Л. Бианки, И.М. Шейман, 1985), в том числе и наши, на беспозвоночных, о том, как наличие ” правшества” или “левшества” связано с проявлением других поведенческих реакций животного, в частности с общедвигательной активностью, способностью к обучению, устойчивостью к загрязнению среды. Все же надо полагать, что механизмы, формирующие асимметрию направления движения, имеют важное биологическое значение, включаясь в осуществлении пространственного анализа и в реализацию двигательных стратегий, т.е. в осуществление важнейших форм ориентационного и поискового поведения. Одностороннее предпочтение может имеет адаптивное значение, обеспечивающее более эффективное исследование и использование животными окружающей среды. У муравьев выбор и закрепление в процессе обучения определенного направления движения является важным элементом стратегии их поведения в многоальтернативной среде, способствующим формированию минимизированных траекторий. Вектор определенного направления движения, входит, по-видимому, как непременный элемент в карту пространства, создаваемую в процессе обучения, способствует анализу пространства по признаку “ правое-левое” (Г.П. Удалова, А.Я. Карась, 1997). Насекомые способны дифференцировать пространство по оси собственного тела, выделяя абстрактные признаки «правый-левый» безотносительно к конкретным ориентирам (Г.А. Мазохин-Поршняков и др., 1987). Выявленная у беспозвоночных, особенно у муравьев, динамичность асимметрии является, как и у позвоночных, особенностью, которая может способствовать эффективному и адекватному изменению поведения в новых условиях.
Сложным является вопрос о связи функциональной моторной асимметрии с асимметрией мозга. По-видимому, там где очевидна морфологическая асимметрия образований головного мозга высока вероятность эндогенной (исходной) функциональной асимметрии (например, у мучного хрущака). Рассматривая анатомо-физиологические предпосылки моторной асимметрии у пчел и ос, предположили, что преимущественный выбор определенного направления движения связан с некими нервными механизмами, т.е с внутренними факторами, но он может формироваться ( или изменяться) под действием многих факторов, в том числе и магнитного поля Земли. (В.М. Карцев и др., 1988).
Нейрофизиологической основой пространственно-моторной асимметрии у беспозвоночных может является и функциональная асимметрия парных образований мозга. У позвоночных по морфологическим, электрофизиологическим и нейрохимическим показателям установлены отчетливые корреляции асимметрии направления движения, как и предпочтения одной из передних конечностей, с асимметрией правых и левых отделов мозга (G. Sherman et al.; S. Glick, D. Ross, 1981; Г.П. Удалова, В.В. Михеев, 1982, 1988; Т. Moran et al., 1984; В.Л. Бианки, 1985; Г.А. Вартанян, Б.И. Климентьев, 1991; Г.П. Удалова, 1996 и др.). На беспозвоночных такого рода исследования в отношении различных форм функциональной асимметрии еще весьма малочисленны (R. Feyereisen, 1985; I. Zakharov, M. Orlova, 1989; Y. Burmistrov, Z. Shuranova, 2000) и проблема эта требует дальнейшей углубленнойразработки. Исследования в этом плане помогут ответить на вопросы о том, почему в ходе эволюции животного мира сформировались билатерально-симметричное строение организмов, а главное, за счет каких механизмов осуществляется дискриминация правой и левой сторон организма и окружающего его пространства.
Литература
Баранюк Г.В., Шишлов С.Б., Яковлев К.А. Изучение зрительного и тактильного анализаторов в различных формах поведения у речного рака // Нейрофизиологические механизмы двигательной активности ракообразных. Л. Изд-во ЛГПИ им. А.И.Герцена. 1983.С.11-21.
Бианки В.Л. Асимметрия мозга животных .Л. Наука.1985. 295 с.
Бианки В.Л., Филиппова Е.Б., Пошивалов В.П. Межполушарная асимметрия двигательной и исследовательской активности у мышей // Журн.высш.нервн.деят. 1982. Т.32. №3. С.442-445.
Бианки В.Л., Шейман И.М. Предпочтение направления движения в Т-образном лабиринте у мучного хрущака // Журн.высш.нервн.деят. 1985. Т. 5. С.988-990
Бианки В.Л., Шейман И.М., Зубина Е.В. Предпочтение направления движения в Т-образном лабиринте у планарий // Журн. высш. нервн. деят. 1990.Т.40. №.1.С.102-107.
Богута К.К. Отношения симметрии в нервной системе низших Bilateria.// Зоол. журн. 1978. Т.57. № 12. С. 1765-1770.
Богута К.К. Некоторые морфологические принципы формирования низкоорганизованных нервных систем в онтогенезе и филогенезе // Зоол.журн.. 1985. Т.64. №11. С. 1605-1613.
Вартанян Г.А., Клеменьев Б.И. Химическая симметрия и асимметрия мозга. Л. Наука. 1991. 150 С.
Глейзер С.И. Функциональная дисимметрия поведения у рыб //Журн. высш. нервн. деят. 1981.Т.31.№.2.С.431-434.
Дашевский Б.А., Карась А.Я..Удалова Г.П. О пластичности поведения муравьев при обучении в многоальтернативном симметричном лабиринте// Журн.высш.нервн.деят. 1989.Т.39. №.1.С.81-89.
Дутов А.А., Анохов С.С. Нейрофизиологическая и фармакологическая характеристики скрытой межкаудатной асимметрии у кошек//Физиол. журн.СССР. 1983. Т.69. N3. С.322-325.
Дутов А.А.. Титова О.Е., Гаврилюк А.Н., Анохов С.С. Выявление скрытой двигательной асимметрии мозга у интактных лабораторных животных // Физиол. журнал СССР. 1985. Т.71.№ 7. С. 925-927.
Захаров А.А., Орлова Т.Н. Поведенческая структура и индивидуальное поведение у муравьев // Сб. Поведение насекомых. М.Наука.1984. С.43-63.
Иванов А.В., Мамкаев Ю.В. Ресничные черви (Turbellaria), их происхождение и эволюция. Л. Наука.1973. 287 С.
Карась А.Я., Удалова Г.П. Пространственно-моторная асимметрия у краба Carcinus meanas при реакции активного избегания // Вестник ЛГУ. 1988.серия 3, №. 2 ( №10 ).С.116-119.
Карась А.Я., Удалова Г.П. Стратегия поведения муравьев при изменении уровня пищевой мотивации // Журн. высш. нервн. деят. 1998.Т.48.№.1.С.91-98.
Карась А.Я., Удалова Г.П. Поведение муравьев в лабиринте при смене пищевой мотивации на защитную // Журн. высш. нервн. деят. 2000. Т.50. №. 4. С. 676-685.
Карась А.Я., Удалова Г.П., Дашевский Б.А. Пространственно-моторная асимметрия при обучении муравьев Myrmica rubra в многоальтернативном лабиринте //М. 1986. 20 С. Деп. в ВИНИТИ, № 6741-В 86.
Карась А.Я., Удалова Г.П., Дашевский Б.А. Некоторые биологические факторы,определяющие функциональную организацию памяти и обучения / на примере изучения поведения насекомых // Сб. Исследование памяти. М. Наука. 1990. С. 119-136.
Карась А.Я., Удалова Г.П., Дашевский Б.А. Ориентация муравьев в лабиринте при поворотах его в горизонтальной плоскости // Сенсорные системы.1995. Т.9. № 2-3. С.50-57.
Карась А.Я., Удалова Г.П.. Загораева Е.В. Роль мотивации при обучении муравьев Myrmica rubra в многоальтернативном лабиринте // Вестник ЛГУ.1986. № 4. C. 43-52.
Карцев В.М., Мазохин-Поршняков Г.А., Семенова С.А. Асимметрия траекторий движения, выбираемая медоносными пчелами и складчатокрылыми осами при поиске корма // Зоол. журн.1988. Т.67. №.8. С.1180-1186.
Клименко Л.Л., Протасова О.В., Конрадов А.А., Фокин В.Ф., Пирузян Л.А. Микроэлементы в нейрофизиологических механизмах формирования межполушарной асимметрии // Изв. РАН. Серия. биол. 1995. № 4.с.422-429.
Мазохин-Поршняков Г.А., Карцев В.М. Изучение последовательности облета насекомыми нескольких равноценных пищевых объектов ( к вопросу о стратегии их визуального поиска ) // Зоол. журн. 1979. Т.58. №.9.С. 1281-1289.
Мазохин-Поршняков Г.А., Карцев В.М., Семёнова С.А., Рабинович А.З. Способность насекомых дифференцировать пространство по признаку справа-слева // Зоол. журн. 1987. Т.66. № 3. С.365-372.
Маркина Н.В., Попова Н.В., Полетаева И.И. Межлинейные различия в поведении мышей, селектированных на большую массу мозга//Журн.высш.нервн.деят. 1999. Т.49. № 1. С. 49-67.
Моренков Э.Д. , Петрова Л.П. Моторная асимметрия у крыс при аудиогенном стрессе. Роль межполушарных отношений // Актуальные вопросы межполушарной асимметрии. Матер. Конференции. М. 2001. С. III-118
Непомнящих В.А., Гремячих В.А. Связь между структурой траектории и асимметрией выбора направления движений у тиляпии Oreochromis mossambicus Peters (Cichlidae) // Журн. общей биологии.1993. Т.54. № 5. C. 619-626.
Рябинская Е.А. Асимметрия направления движения у крыс линии Вистар и . Крушинского-Молодкиной в радиальном лабиринте// Журн. высш.нервн деят. 1982. Т.32. №. 3. С.566-568.
Рябинская Е.А.,Валуйская Т.С. Асимметрия направления движения как тактика пищевого поведения у крыс // Журн.высш.нервн.деят. 1983. Т.33.№.4. С. 654. 661.
Серавин Л.Н., Карпенко А.А. Особенности ориентировки беспозвоночных в трехмерном пространстве// Зоол. журн. 1987. Т.66, вып. 9, С. 1285-1292.
Тхоржевский В.В. Изучение реакции выбора сторон у мышей // Журн.высш.нервн.деят. 1973. Т.23.№.3. С.659-661.
Удалова Г.П. Участие правого и левого полушарий в реализации . лабиринтного навыка у мышей-самцов линии ВАLB/с.// Журн. высш. нервн. деят. 1996. Т.46. № 1. С.84-92
Удалова Г.П., Жуковская М.И., Карась А.Я. Способность муравьев к множественным переделкам лабиринтного навыка // Журн. высш. нервн. деят. 1991. Т.41. № 6. С. 1154-1162.
Удалова Г.П.. Жуковская М.И., Карась А.Я. Пространственно-моторная асимметрия у муравьев при множественных переделках лабиринтного навыка // Вестник СПбГУ. 1992. Сер. 3. №.1 ( 3 ) .С. 67-75.
Удалова Г.П., Карась А.Я. Асимметрия направления движения у муравьев Myrmica rubra при обучении в лабиринте // Журн.высш.нервн.деят. 1985.Т.35. №.2.С.377-379.
Удалова Г.П., Карась А.Я. Асимметрия направления движения у муравьев Myrmica rubra при обучении в лабиринте в условиях пищевой мотивации // Журн. высш. нервн. деят. 1986. Т.36. №. 4. С. 707-714..
Удалова Г.П., Карась А.Я. Асимметрия направления движения у муравья Myrmica rubra // Журн. эволюц. биохимии и физиол. 1989а. Т. 25. № 1. С. 27-37.
Удалова Г.П., Карась А.Я. Пространственно-моторная асимметрия у муравьев / Нейробиология церебральной латерализации.Л.ЛГУ. 1989 б. С.173-198.
Удалова Г.П., Карась А.Я. Значение различных сенсорных стимулов для ориентации муравьев // Вестник СПбГУ, 1997а. сер.3.№.2.С.62-68.
Удалова Г.П., Карась А.Я. Анализ пространства по признаку “правое-левое” у муравьев Myrmica rubra / Матер. Международных коллоквиумов по общественным насекомым. Proc. Int. Coll. Social. Insects. 1997 б . Vol.3-4. .P. 287-292.
Удалова Г.П., Карась А.Я. Индивидуальные особенности модификации лабиринтного навыка при изменении вида и уровня мотивации у муравьев Myrmica rubra // Успехи современной биологии. 1999. Т. 119. № 3. С. 233-242.
Удалова Г.П., Карась А.Я.. Жуковская М.И. Асимметрия направления движения у гаммарусов ( Gammarus oceanicus ) в тесте открытого поля // Журн. высш. нервн. деят.1990 а. Т.40. № 1. С. 93-101.
Удалова Г.П., Карась А.Я., Жуковская М.И. Влияние нефтяного загрязнения на поведение бокоплавов Gammarus oceanicus в тесте “открытое поле“// Физиология и токсикология гидробионтов. Ярославль.ЯГУ.1990 б. С.56-64.-
Удалова Г.П., Кашина И.А. Полушарная асимметрия двигательно-пищевого поведения мышей в многоальтернативном симметричном лабиринте // Журн.высш.нервн.деят. 1987. Т.34. №.4. С.703-710.
Удалова Г.П., Михеев В.В. Роль функциональной межполушарной асимметрии в формировании предпочтения направления движения у крыс// Журн. высш. нервн. деят. 1982. Т.32. №.4. С.633-641.
Удалова Г.П.. Михеев В.В. Об участии полушарий в формировании пространственно-моторной асимметрии при зрительном распознавании у крыс // Журн. высш. нервн. деят., 1988. Т. 38. №. 3. С. 467-474.
Цыганов В.В. Координация моноаминергических педальных нейронов у пресноводных улиток // Российский .физиол.журнал. 2000. Т. 86. № 4 .С.369-378.
Bretz D.D. , Dimock R.V. Behaviorally important haracteristics of the mucous trail of the marine gastropod Hyanossa obsoleta (Say) // J. Exp. Biol.and Ecol. 1983,Vol.71. № 2, P.181-191.
Brihschneider F. Uber die Gehirne der Kuchensehabe und des Mehlkafers //Naturwissenschaften, 1914. Bd. 52. №2.S 269.
Burmistrow Yu., Shuranova Zh. Field potentials generated by symmetrical ventilatory appendages in free moving crayfish // Simpler nervous systems. Abstracts of 6th East European conference of the international society for invertebrate neurolology. Moscow-Pushchino. 2000. P.38.
Chapounthier G., Pallaud B., Ungerer A. Relations entre deux reactions des planaires face a une discrimination droite-gauche //C.r. Acad. Sci. 1968. D 266, Vol. 9, 905-907.
Cheverton J. Bumblbees may use a suboptimal arbitrary handedness to solve difficalt foraging decisions // Anim. Behav. 1982. Vol.30. № 3. P.934-935.
Denenberg V. Lateralization of function in rats // Amer. J. Physiology. 1983. V. 245. P.
Feyereisen R. Regulation of juvenile hormone fiter: syntesis // Comprehensive insect physiology, biochemistry and pharmacology. Pergamon Press. 1985. Vol. 7. P. 391-422.
Giehrl T., Distel H. Asymmetric distribution of side preference in hamsters can be reversed by lesions of the caudate nucleus // Behav. Brain Res. 1980.Vol. 1. № 2. P. 187-1962.
Glick S.D., Cox R. Differential effects of unilateral and bilateral caudate lesions on side preferences and turning behavior in rats // J. Compar. and Psychol. 1976. Vol. 90. P 528-538.
Glick S.D., Ross D.A. Right-sides population bias and lateralization of activity in normal rats // Brain Res. 1981. Vol. 205. P. 222-225.
Glick S. D., Ross D.A., Hough L.B. Lateral asymmetry of neurotransmitters in human brain // Brain Res. 1982. Vol. 234. № 1. P. 53-63.
Glick S.D., Weaner L.M., Meibach R.C. Amphetamine-induced rotation in normal cats // Brain Res. 1981. Vol. 208. P. 227-229
Glick S., Yerussi T., Zimmerberg B. Behavioral and neuropharmacological correlates of nigrostriatal asymmetry in rats//In: Lateralization in the nervous system. Ed. S. Harnad. NY. 1977. P. 213-249.
Hoffman J. Running strategy and turning preference of rats in Y-maze // Acta Physiol Acad.Sci. Hung. 1980.Vol. 56. № 1. P. 114
Korczyn A.D., Eshel Y. Dopaminergic and non-dopaminergic circling activity of mice // Neuroscience. 1979. Vol.4. № 8. P. 1085-1088.
Kubos K., Robinson R. Cortical undercuts in the rat produce asymmetrical behavioral response without altering chatecholamine concentrations // Exper. Neurology. 1984. Vol. 83. P. 646-653.
Moran T. H., Sanberg P.K. , Kubos K.L. et al. Аsymmetrical effects of unilateral cortical section lesions // Behav. Neuroscience. 1984. Vol. 98. № 4. P.747-752.
Nottebohm F. Origin and mechanisms in establisment of cerebral djminance//Handbook of behavioral neurobiology. Ed. M.Gazzaniga. N.Y. L. Plenum Press. 1979. V.2. P. 295-344
Putnam C. The non-random behaviour of Aleochara bilineata Gyll. (Coleptera; Stophylinidae ) in a Y-maze with neither reward nor punishment in either arm//Animal behaviour. 1962. Vol.10. № 182. P .118-125.
Ross D.A., Glick S.D. Lateralized effects of bilateral frontal cortex lesions in rats //Brain Res. 1981. Vol. 210. P. 379-382.
Schmid-Hempel P. The importance of handling time for the flight directionality in bees // Ecol. and Sociobiol. 1984. Vol. 15. № 4. P. 303-309.
Sherman G.F., Garbanati J.A., Rosen G., Yutzey D., Denenberg V. Brain and behavioral asymmetries for spatial preference in rats // Brain Res. 1980. Vol. 192.№ 1. P. 61-67
Wilson D.M. Inherent asymmetry and reflex modulation of the locust flight pattern // J. Exp. Biol. 1968. Vol. 48. P. 631-641.
Witt P.H. Lateralization and the invertebrate nervous system.Discussion // In: Evolution and lateralization of the brain. Eds. E.J.Dimond, D.A.Blizard.Ann.N.Y.Acad.Sci..1977.Vol.299. P.72.
Zakharov I., Orlova M. Gfad-containing neurons take part in the control of mating behavior // Simpler neurous systems. Absracts of the 5th East European conference of the international society for invertebrate neurobiology. Moscow. 1989. P.96.