ЯВЛЕНИЯ ФУНКЦИОНАЛЬНОЙ АСИММЕТРИИ У ПЛАНАРИЙ
И.М. Шейман, Е.В. Зубина, В.Л. Бианки
Институт Биологической физики АН СССР, Пущино;
Биологический Институт СПбУ, Санкт-Петербург
В течение долгого времени бытовало убеждение, которое не исключено и сейчас, что асимметричная функциональная организация мозга – свойство высоко организованных существ. Постепенно пробивают себе дорогу сведения об асимметрических явлениях среди разных животных. Беспозвоночные животные зачастую остаются вне поля зрения данной проблемы. Вместе с тем описан ряд успешных попыток выявления у них функциональной асимметрии мозга. Такие исследования проведены на пчелах, жуках, муравьях (В.Л. Бианки, 1985), моллюсках (Н.Б. Салимова, И. Милошевич, Р.М. Салимов, 1984), раках (Г.П. Удалова, А.Я. Карась, М.И. Жуковская , 1990), крабах (А.Я. Карась, Г.П. Удалова, 1988), муравьях (Г.П. Удалова, А.Я. Карась, 1985, 1986) В перечисленных работах изучалась пространственно-моторная асимметрия мозга. Возникает естественный вопрос, в какой мере феномен функциональной асимметрии связан с эволюционным развитием нервной системы.
Центральная нервная система в филогенетическом ряду впервые возникла у плоских червей. Ключевая позиция плоских червей в эволюционном ряду определяется концентрацией нервных элементов в ортогон и выделением головного конца нервной системы в парный ганглий (В.Н. Беклемишев, 1964; К.К. Богута,1978), а также появлением билатерально - симметрической организации тела. В этой связи исследование функциональной симметрии и асимметрии у плоских червей представляет значительный интерес.
Некоторых плоских червей – планарий - отличает исключительная способность к регенерации любой части тела после повреждения, перерезки и деления (бесполого размножения). Процесс регенерации у планарий зависит от наличия центральной нервной системы и регулируется ею (И.М. Шейман, 1984).
Все сказанное выше явилось предпосылкой для исследования у планарий особенностей пространственной ориентации в Т-образном лабиринте и изучения у них характера регенерации в симметричных частях тела.
Использовались пресноводные планарии Dugesia tigrina (предположительно половая и бесполая расы), Dugesia lugubris, Ijimia tenuis и Bdellocephala punctata, являющиеся представителями отряда Seriata, класса Turbellaria (ресничные черви), типа Plathelmintes (плоские черви). Эти животные ведут образ жизни свободных хищников.
Эксперименты в Т-образном лабиринте проводились в 1985-1988 гг. на 1768 планариях в г. Пущино-на-Оке. Опыт на 1353 планариях D. tigrina бесполой расы были проведены летом 1985 г., летом, осенью и зимой 1986 г. и летом и зимой 1987г.; опты на 100 планариях D. tigrina половой расы – летом 1987 г.; опыты на 100 планариях D. lugubris – зимой 1988 г.; опыты на 100 планариях Ij tenuis летом 1985 и 1987 гг.; опыты на 55 планариях Bd. punctata - летом 1988 г. Планарии Ij. tenuis и Bd. punctata были собраны в старицах Оки и содержались затем в лабораторных условиях. D. lugubris были доставлены из прудов Петергофа D. tigrina половой расы были взяты из аквариума Института Биофизики а бесполая раса D. tigrina является лабораторной и также разведена в аквариумах Института.
В работе использованы главным образом планарии D. tigrina бесполой расы.
Планарии жили в затемненных аквариумах, заполненных водопроводной водой, на 1/3 разбавленной дистиллированной водой. Вода не продувалась. Температура поддерживалась на уровне 20-25°С. Планарий кормили 1 раз в неделю мотылем, и в эксперименте использовали не раньше, чем через 2 сут после кормления.
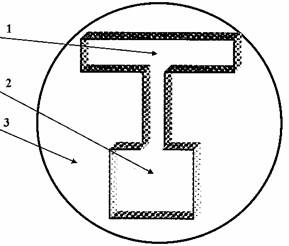
Применяли методику свободного выбора направления в Т-образном лабиринте. Использовали плексигласовый лабиринт, состоящий из коридоров длиной 5 см глубиной 5 мм, шириной 6 мм и стартовой камеры размером 2х2 см той же глубины. Боковые коридоры свободно открывались наружу, а стартовый – в стартовую камеру. Лабиринт помещали в кристаллизатор с водой, которая заполняла лабиринт на 2/3 глубины (рис.1).
Рис. 1. Т – образный лабиринт.
1 – свободный выбор направления; 2 – стартовая камера; 3 - кристаллизатор с водой.
Опыт проводили параллельно в трех лабиринтах. Освещенность была равномерной и постоянно контролировалась. Температура воздуха в лаборатории колебалась в пределах 20-25°С.
Во время опыта планарий рассаживали по одной в пронумерованные стаканчики. Манипуляции проводили с помощью пипетки. Опыт состоял из 50 проб (5 серий по 10 проб в каждой). Планарию помещали в стартовый коридор головным концом в направлении развилки. После выбора одного из направлений планарию вновь переносили в стартовый коридор, и так 10 проб подряд. Затем планарию возвращали в затененный стаканчик и заменяли следующей. В течение дня в каждом лабиринте попеременно «работали» 5 планарий. После того, как заканчивали серию на 5 животных, лабиринт протирали ватой для удаления слизи и начинали следующую серию. Таким образом, каждая планария отдыхала между сериями примерно 1 ч. В течение одного дня в опытах участвовали 15 планарий. Отмечали направление каждого сделанного планарией выбора. Суммировали число правых и левых поворотов по сериям и в целом за весь опыт.
Так как при своем движении планария выделяет слизь, неоднократное использование лабиринта для одного и того же или разных животных вызывало опасение о возможной сигнальной роли слизи. Для элиминации этого предположения на 45 планариях проводили специальные контрольные опыты, в которых после каждой пробы лабиринт протирали спиртом и тщательно промывали водой. Результаты контрольной серии экспериментов полностью совпадали с основными опытами.
В опытах по регенерации планарий рассекали по продольной оси тела и ампутировали у них глотку (рис.2).
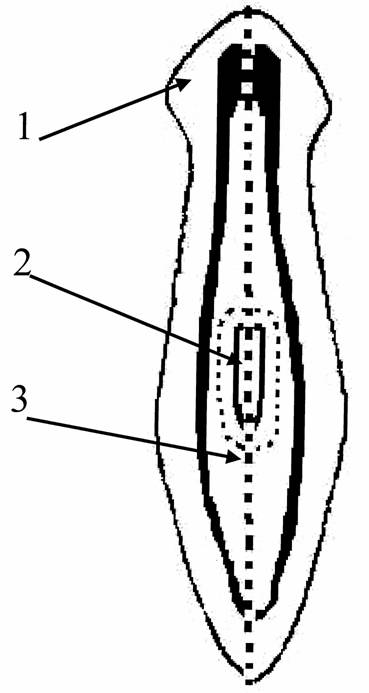
Рис. 2. Планария и схема операций.
1 – центральная нервная система планарии; 2 – глотка; 3 – пунктир обозначает направление разреза и удаление глотки.
Операцию проводили глазным скальпелем под бинокулярным микроскопом. Правые и левые фрагменты тела размещали отдельно по 10 в стаканчики с 50 мл воды. Показателем регенерации служило функциональное восстановление глотки, о которой судили по появлению пищедобывательной реакции глотки. С этой целью в последующие после операции дни регенерантам предлагали мотыль. Отмечали первый день, когда каждая планария начинала есть.
Для анализа регенерации симметричных половинок тела исследовали влияние на этот процесс нейрорегуляторных пептидов, обладающих морфогенетической активностью: морфоген гидры, или головной активатор гидры (HHA), люлиберин (LHRH), меланостатин (MST), тиролиберин (TRH), даларгин (DAL) – все в концентрации 10-9 М, и вазопрессин (VP) – в концентрации 10-7 М.
После операции планарий помещали в соответствующие растворы пептидов по 10 в 50 мл. Каждая исследованная группа состояла из 50 животных. Опыты поставлены на планариях D. tigrina.
Данные всех экспериментов обрабатывали статистически с помощью непараметрического критерия знаков, t-критерия Стьюдента, критерия «c-квадрат» и однофакторного дисперсионного анализа.
В первую очередь было изучено предпочтение планариями направления движения в Т-образном лабиринте в условиях свободного выбора при проведении 50 последовательных проб. Были использованы 721 особь D. tigrina бесполой расы, 100 особей D. tigrina половой расы, 160 особей Ij. tenuis и 55 особей Bd. punctata – всего 1136 животных. По характеру поведения в лабиринте выделяли следующие группы планарий: RL-формы (или «симметрики»), не обнаружившие достоверного предпочтения одного какого-либо направления движения, и R+ L-формы (или «асимметрики») – группа, в которую входили R-формы (или «правши») предпочитающие правый поворот, и L-формы (или «левши»), предпочитающие левый поворот. Наличие симметрии или асимметрии направления движения идентифицировали статистически у каждого животного отдельно на основании его поведения в 50 последовательных пробах с помощью критерия знаков.
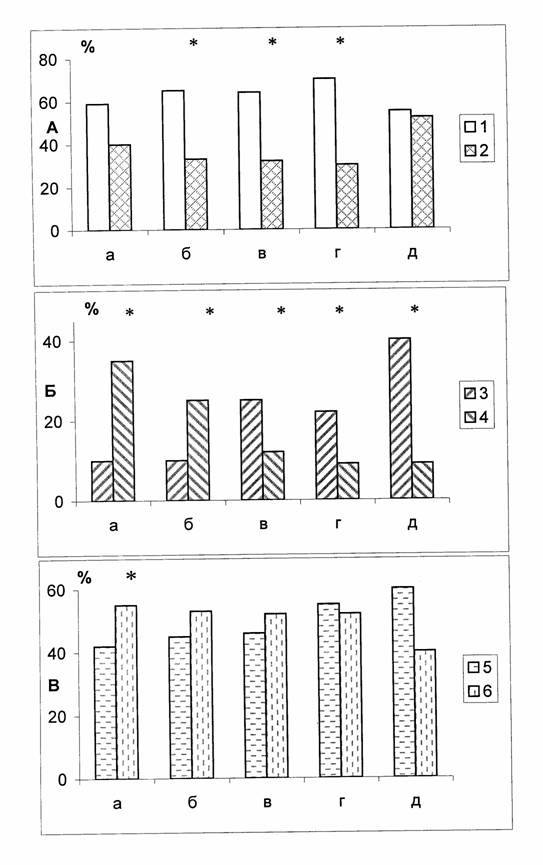
Рис.3. Распределение планарий по типу предпочтения направления поворота в лабиринте.
А – симметрики RL - формы (1) и асимметрики (2) R+L - формы. Б – левши L - формы (3) и правщи (4) R - формы. В – общее количество левых (5) и правых (6) поворотов а – D. tigrina (бесполая раса); б - D. tigrina (половая раса) ; в - D. lugubris; г - Ij. Tenuis; д - Bd. Punctata. – * достоверность различий при р< 0.05, ** достоверность различий при р <0.01.
Видно (рис.3, А), что все исследованные животные могли быть разделены на две неодинаковые по величине группы «симметриков» и «асимметриков», причем группа «симметриков» была большей и состояла в среднем из 60 % исследованной выборки, а группа «асимметриков» – меньшей и включала в себя лишь 40 % выборки. Преобладание RL-форм над R+L-формами для видов D. tigrina (половая раса), D. lugubris и Ij. tenuis было достоверным (р<0,05), а для бесполой расы D. tigrina и Bd. punctata выражалось в виде тенденции (p>0,05).
Группа «асимметриков» по своему составу тоже оказалась неоднородной (рис.3, Б). У вида D. tigrina (бесполая и половая расы) преобладали «правши», составлявшие 23-32% от общей выборки, а «левши» составляли лишь 10% (р<0,05). С другой стороны, у видов D. lugubris, Ij. tenuis, Bd. punctata «левши» преобладали над «правшами», составляя соответственно 22-40 и 9-11% (p<0,05<0,01).
Таким образом, оказалось, что у планарий симметричные формы поведения в Т-образном лабиринте преобладают над асимметричными, а «асимметрики» подразделяются на две группы – виды с превалированием правых или левых выборов направления поворота.
Для последующего анализа у тех же животных сравнивали общее количество поворотов, совершенных в ту и другую сторону в популяции в целом (рис.3, В). Оценка соответствующих результатов обнаружила достоверное (p<0,05) преобладание правосторонних реакций у бесполой расы D. tigrina над левосторонними (56 и 44 % соответственно). У других исследованных видов, представленных выборками меньшей величины, наблюдалась лишь тенденция к преобладанию количества правых или левых поворотов. При этом можно было отметить соответствие между количеством R- и L- форм в популяции и тенденциями к преобладанию числа поворотов вправо и влево.
Представляет интерес соотношение направления первого и последующих поворотов планарий в Т-образном лабиринте. Этот показатель в известной мере дает представление об устойчивости выявленного предпочтения. Данный вопрос исследовали на 721 планарии D. tigrina бесполой расы. Из таблицы видно, что общее количество первичных правых и левых поворотов в исследованной популяции различалось незначительно, хотя и наблюдалась известная тенденция в сторону преобладания правых поворотов, которая была сильнее выражена у «асимметриков» по сравнению с «симметриками». Анализ направления первого поворота у R- и L- форм обнаружил следующую закономерность: из общего числа R-форм примерно у 2/3 первая реакция была правосторонней, а у 1/3 – левосторонней. Точно так же примерно у 2/3 L-форм первая реакция была левосторонней, а 1/3 – правосторонней.
Таблица
|
Группы планарий |
Направление первого поворота |
|
|
Вправо |
Влево |
|
|
R-формы («правши») |
153 |
48 |
|
L-формы («левши») |
24 |
71 |
|
Всего |
177 |
119 |
|
RL-формы («асимметрики») |
220 |
205 |
|
Всего |
397 |
324 |
Несколько более полное представление о прочности «поворотных тенденций» у планарий дает сопоставление направления движения не в 50 пробах, как рассматривалось выше, а в 250. В этих экспериментах участвовали 30 планарий бесполой расы D. tigrina. Опыты проводили через день в течение 10 суток. По результатам первого опыта планарии разделялись на три группы: «симметриков» (RL-формы), «правшей» (R-формы) и «левшей» (L-формы). Во всех опытах подсчитывали количество правых поворотов для каждой выделенной группы животных (рис.4).
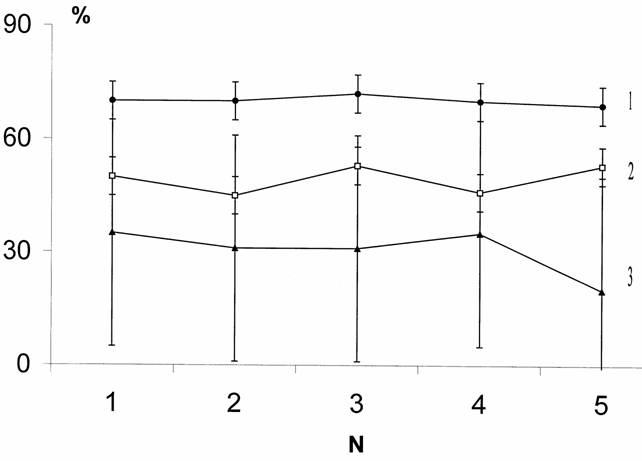 Рис.4. Стабильность предпочтения направления движения у
планарий.
Рис.4. Стабильность предпочтения направления движения у
планарий.
По оси абсцисс – номера опытов, по оси ординат - повороты вправо, проценты. 1 – R - формы (правши) , 2 – RL - формы (симметрики), 3 - L - формы (левши).
В течение пяти опытов (т.е. на протяжении 10 дней) количество правосторонних поворотов для каждой группы было практически постоянным, а данные разных групп планарий значительно различались между собой. Используя индивидуальные показатели каждого животного, анализировали прочность уровня выбора правого поворота. Оценивали дисперсию реакций одного животного в течение всего периода опытов относительно дисперсии реакции разных животных в течение одного дня. Было показано, что стабильность уровня выбора правого поворота характерна не только для групп в целом, но и для каждого животного в отдельности. Результаты по разным планариям в один день различались достоверно больше, чем данные одного и того же животного на протяжении 10 дней (p<0,01).
В ходе экспериментов контролировали постоянство внешних условий. Поскольку известно влияние сезонных изменений на поведение планарий, специальное внимание уделяли изучению влияния времени года на «поворотные тенденции». С этой целью были проанализированы данные основной серии опытов, проведенной на 721 планарии D. tigrina бесполой расы, из которых 497 особей исследовали летом, 180 – зимой и 44 осенью (рис.5).
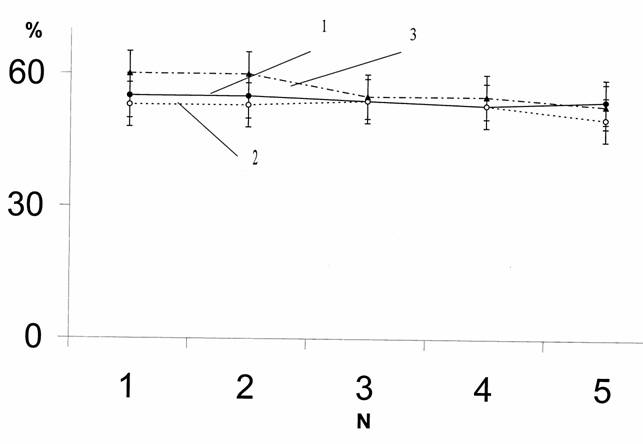
Рис.5. Влияние времени года на поворотные тенденции у планарий.
1 – лето, 2 – зима, 3 – осень. Обозначения те же, что на рис.4.
Средние данные по количеству поворотов вправо из 50 проб оценивались по t-критерию Стьюдента. Планарии поворачивали вправо в среднем: 27,7 раза летом, 27,3 раза зимой и 29,1 раза осенью (p>0,05). Таким образом, полученные данные не выявили сезонных изменений в характере выбора направления поворота у планарий, что опять-таки свидетельствует о стойкости этого показателя.
В поисках возможных факторов, обусловливающих выбор планариями предпочитаемого направления движения, было проанализировано и влияние ориентации лабиринта. В различных сериях опытов перед началом движения в лабиринте планарии были направлены головным концом тела в разные стороны. Эксперименты проводили на 721 планарии D. tigrina бесполой расы. При северной ориентации исследовано 423 планарии, при южной – 148, при восточной и западной – по 75 планарий.
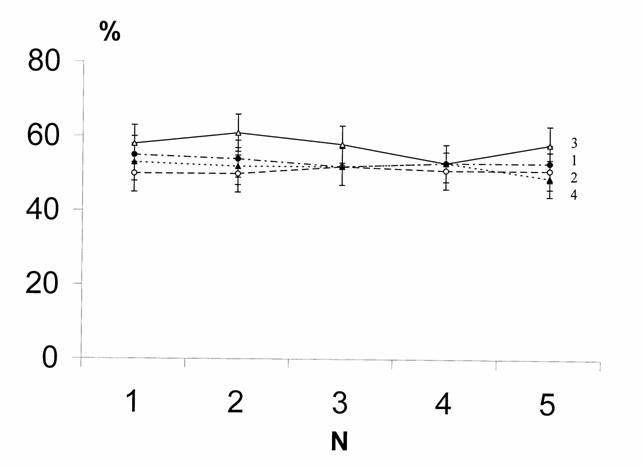
Рис.6. Влияние ориентации лабиринта на поворотные тенденции у планарий.
1 – север, 2 – юг, 3 – запад, 4 – восток. Обозначения те же, что на рис. 4.
Можно видеть (рис.6), что ориентация лабиринта значимо не влияла на предпочитаемое направление движения. При северной ориентации был сделан в среднем 28,1 правого поворота, при южной – 26,1, при западной – 29,9, и при восточной – 26,0. Эти различия были статистически недостоверными (p>0,05).
В каждом опыте на оперированных планариях использовали по 50 животных. Опыты повторяли 2-3 раза. Регенерантов содержали по 10 в стаканчиках с водой или раствором пептида, отдельно правые и левые фрагменты.
В первой серии опытов на разрезанных вдоль планариях было показано, что пищедобывательная реакция глотки восстанавливалась у регенерантов в течение 5-8 дней после операции При этом в средних сроках ее восстановления между правыми и левыми фрагментами не было достоверных отличий (р>0.05) (рис.8, контроль).
Функциональное восстановление глотки определяется не только регенерацией ее структуры, но также зависит от регенерации нервного аппарата. Опыты на правых и левых фрагментах тела планарий показали, что регенерация в целом произошла у них одинаково. Углубленный анализ этих данных был произведен на основе биохимического подхода, а именно с использованием регуляторных нейропептидов.
Ранее было установлено, что некоторые нейропептиды обладают морфогенетической функцией (И.М. Шейман, Х.П. Тирас, Э.Ф. Балобанова, 1989) и действуют на регенерацию глотки у целых планарий как стимуляторы процесса, в то время как другие пептиды его тормозят (Н.Д. Крещенко, И.М. Шейман 1994).
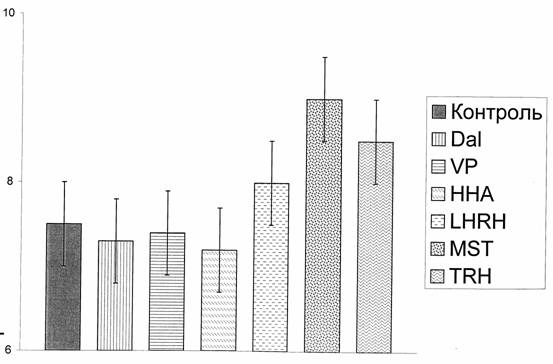
Рис. 7. Время восстановления пищедобывательной реакции глотки у планарий (названия нейропептидов см. в тексте, в разделе методика). По оси ординат – время регенерации.
На рис.7 видно, что к группе стимуляторов восстановления пищедобывательной реакции глотки отнесены морфоген гидры, даларгин и вазопрессин, а к группе ингибиторов -.люлиберин, меланостатин и тирoлиберин.
При действии перечисленных пептидов на регенерацию глотки и восстановление ее функции у правых и левых половинок тела были получены результаты, подтверждающие данные, выявленные на целых планариях. Однако эффект, полученный на разных фрагментах планарий, отличался как при действии стимуляторов, так и при действии ингибиторов регенерации. Так, морфоген гидры, даларгин и вазопрессин достоверно сокращали время восстановления функции глотки у правых фрагментов планарий (р<0.05), т.е. действовали как стимуляторы. В то же время даларгин и вазопрессин стимулировали левые фрагменты незначительно (Р>.05). Обратная картина наблюдалась при действии ингибиторов регенерации. Люлиберин, тиролиберин и меланостатин достоверно (р<0.05) задерживали время восстановления функции глотки у левых половинок планарий. Действие меланостатина и тиролиберина на правые половинки было слабее выражено, хотя и имело достоверный характер, а действие люлиберина на правые половинки было незначительным. Все эти данные приведены на рис 8.
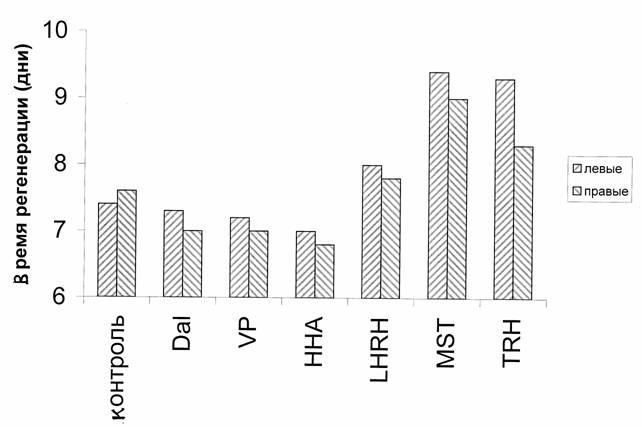
Рис.8. Время восстановления пищедобывательной реакции глотки при регенерации левых и правых фрагментов планарий.
Кроме того их иллюстрирует схема, приведенная на рис.9.
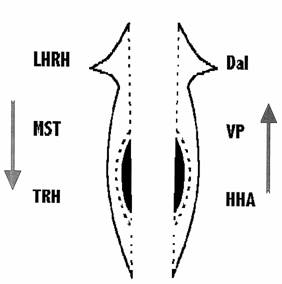
Рис.9 Схема действия нейропептидов на скорость восстановления пищедобывательной функции глотки у правых и левых фрагментов планарий. Стрелки указывают на стимулирующее (вверх) или тормозящее (вниз) действие.
Плоские черви, являются важнейшим объектом для изучения латерализации функций, так как у них впервые в эволюционном ряду возникает билатеральная симметрия в структурной организации тела. Это явилось одним из поводов для утверждения, что беспозвоночные в подавляющем большинстве устроены и функционируют на основе билатеральной симметрии (А.В. Иванов, 1968). Высказывалось также мнение, что плоские черви не могут различать «правое» и «левое», так как они обладают выраженной билатеральной симметрией, а некоторое различение развивается лишь в ходе эволюции по мере прогрессирующей утраты степени симметрии тела (А.В. Иванов, Ю.В. Мамкаев,1973). Вместе с тем проблема появления асимметрии функций у беспозвоночных поднималась ранее в связи с филогенезом нервной системы (W.D. Chapple, 1977, G.S. Berkowitz, R.D. Tschirgi, 1984).
Проведенные нами исследования показали, что у большинства планарий при предпочтении направления движения «асимметриками» оказались 1/3-1/2 популяции. У более высоко организованных животных «асимметрики», как правило, представлены меньшинством, и часто очень незначительным (В.Л. Бианки,1985).
Вместе с тем, обнаруженная у планарий форма функциональной асимметрии в направлении движения противоречит приведенным утверждениям, не основанным на прямых экспериментах. Описанные данные показывают, что функциональная асимметрия в филогенетическом ряду возникает, вероятно, одновременно с билатеральной симметрией.
Наиболее существенным из полученных фактов является тот, что у всех исследованных видов планарий «асимметрики» разделялись на две неравные группы «правшей» и «левшей», причем различия между ними были статистически достоверными. Эта асимметрия была относительно стабильной, и в частности не зависела от времени года и ориентации по сторонам света. У D. tigrina (бесполая и половая расы) преобладали «правши», а у D. lugubris, Ij.tenuis, Bd. punctata – «левши». Следовательно, направление асимметрии могло различаться не только у различных видов разных родов, но и у одного и того же рода (Dugesia). Привлекает внимание то обстоятельство, что у лабораторного вида (D. tigrina) асимметрия правосторонняя, а у остальных видов, взятых из природы, - левосторонняя.
Приведенные факты наводят на мысль о том, что уже у планарий – наиболее примитивных билатерально-симметричных животных – мы встречаемся с какой-то загадочной формой видовой функциональной асимметрии. В самом деле, во-первых, преобладание у них того или другого знака асимметрии является видоспецифическим признаком для асимметричной части популяции; во-вторых, у всей исследованной популяции в целом могут преобладать однонаправленные «поворотные тенденции». Вместе с тем отношение асимметричных особей вида к их общему количеству является еще относительно небольшим, что позволяет характеризовать описанную выше форму первичной асимметрии как примитивную.
Наряду с выявленной формой частично скрытой функциональной асимметрии в направлении движения, обнаружилась также замаскированная асимметрия в характеристике нервной системы планарий, выявленная в морфогенетическом процессе. Регенерация двух симметричных фрагментов тела, правой и левой половинок планарий, исследованная на основе восстановления пищедобывательной функции глотки, происходила без видимых различий. Пищедобывательная реакция восстанавливалась в одни и те же сроки у правых и левых регенерантов. Ранее было установлено, что ее восстановление определяется как морфологической регенерацией глотки, так и функционированием нервной системы планарий.
Принято считать, что функциональная асимметрия мозга - явление присущее всем позвоночным животным, хотя ряд работ, выполненных на беспозвоночных животных позволил выявить у них также функциональную асимметрию мозга. Попытка обнаружить ее у планарий, у которых имеется первая в филогенетическом плане центральная нервная система, не менее важна, чем ее связь с билатеральной организацией тела. Первый экспериментальный подход к задаче, казалось бы, не выявил различий между восстановлением функции в двух симметричных частях тела. Однако применение специальной группы веществ, регуляторных нейропептидов, в опытах с восстановлением пищедобывательной реакции у регенерантов изменил картину. Оказалось, что под влиянием нейропептидов реакция в правых и левых регенерантах восстанавливалась с различной скоростью. Все вещества в той или иной мере оказывали влияние на скорость восстановления реакции. При этом наблюдалось различное их действие на правые и левые фрагменты. Интересно, что морфогенетические стимуляторы (морфоген гидры, даларгин и вазопрессин) оказывали более сильное действие на правые половинки тела, а морфогенетические ингибиторы – на левые. Эти данные указывают на различную химическую характеристику нервных образований в симметричных половинках тела.
Нейрохимическая латерализация центральной нервной системы позвоночных животных известна (В.К. Луценко, М.Ю. Карганов, 1985, Г.А. Вартанян, Б.И. Клементьев, 1988). Показано также, что нейрохимическая асимметрия может быть усилена введением веществ (B. Zimmerberg, S. Glick, T. Jerussi, 1974). Представлкенные результаты опытов вскрыли биохимическую асимметрию центральной нервной системы уже на уровне плоских червей.
Таким образом, наличие латерализации при выборе направления движения вскрыло появление зачаточной функциональной асимметрии одновременно с возникновением билатеральной симметрии, а влияние нейропептидов на восстановление пищедобывательной реакции симметричных фрагментов планарий в ходе регенерации обнаружило, что асимметрическая химическая характеристика нервной системы возникает одновременно с появлением центральной нервной системы как таковой.
Беклемишев В.Н. Основы сравнительной анатомии беспозвоночных. – 1964 – Т.1-2. М.
Бианки В.Л. Асимметрия мозга животных. – 1985 -. Л., Наука, С.295
Богута К.К. Отношение симметрии в нервной системе низших Bilateria // Зоол. журн. – 1978 – Т.57, № 12, С. 1765 – 1770.
Вартанян Г.А., Клементьев Б.И. Проблема химической асимметрии мозга // Физиология человека. – 1988. – Т. 14, № 2, С. 297 – 313.
Иванов А.В. Происхождение многоклеточных животных. – 1968 Л., Наука, С.287.
Иванов А.В., Мамкаев Ю.В. Ресничные черви, их происхождение и эволюция. – 1973 - Л., Наука, С. 245.
Карась А.Я., Удалова Г.П Пространственно – моторная асимметрия у краба. Carcinus meanas при реакции активного избегания // Вестник ЛГУ. – 1988 – Сер.3, № 2, С.116 - 119.
Крещенко Н.Д., Шейман И.М. Регенерация глотки у планарий. Действие нейропептидов. // Онтогенез.- 1994 - Т. 25, № 5, С.350 – 356.
Луценко В.К., Карганов М.Ю. Биохимическая асимметрия мозга //Биохимия. – 1985 – Т. 4, № 2.С.197 – 213.
Салимова Н.Б., Милошевич И., Салимов Р.М. Действие 5,6 – окситриптамина на поведение в лабиринте улитки. // Ж. высш. нервн. деят. – 1984 – Т.34, № 5, С.941 – 947.
Удалова Г.П., Карась А.Я. Асимметрия направления движения у муравьев.Murmica rubra при пищевой мотивации // Ж. высш. нервн. деят.. –1986 - Т.36, № 4, С.707 – 714.
Удалова Г.П., Карась А.Я., Жуковская М.И. Асимметрия направления движения у гаммарусов Gammarus oceanicus в тесте открытого поля. // Ж. высш. нервн. деят. – 1990 - Т. 40, № 1, С.93 – 100.
Шейман И.М. Морфогенетически активные вещества и их адаптивная роль. – 1984 – М, Наука, С. 187.
Шейман И.М., Тирас Х.П., Балобанова Э.Ф. Морфогенетическая функция нейропептидов // Физиолог. Журнал им. Сеченова - 1989 – Т. 75, № 5, С.619-626..
Berkowitz G.S., Tschirgi R..D. On the phylogeny of asymmetry and special discrimination // J. Theoret. Biol. –1984 – V.106, P. 495 – 501.
Chapple W.D. Role of the asymmetry in yhe functioning of the invertebrates nervous systems . In : Lateralization in the nervous system.
Harnad S et al eds. – 1977 – London, Academic press, P. 3-22.
Zimmerberg B., Glick S.D., Jerussi T.P. Neurochemical correlates of a spatial preferance in rats // Science – 1974 – V. 185, N 4151, P. 623 – 625
Gerschwind N., Galaburda A.M. Cerebral lateralization. Biological mechanismes, association and pathologicaal. // Arch. Neurol. –1985 – V.42, N 5, P. 428-459.