РАЗЛИЧИЯ ПОЛУШАРИЙ В ОБРАБОТКЕ ЗРИТИЛЬНОЙ ИНФОРМАЦИИ И ОПОЗНАНИИ ЗРИТЕЛЬНЫХ ОБРАЗОВ
Л.И. Леушина, А.А. Невская
Институт физиологии им. И.П. Павлова РАН
Институт раннего вмешательства, Санкт-Петербург
Исследования, проведенные на лицах с расщеплённым мозгом (M.S. Gazzaniga et al., 1965; J. Levy et al., 1972; R.W. Sperry et al., 1979 и др.), когда есть возможность направлять зрительный стимул только в одно из полушарий и вся обработка зрительной информации происходит только в одной половине мозга, показали, что как левое, так и правое полушарие способно узнавать видимые предметы и даже сложные сцены. Левое полушарие при этом может давать вербальный отчёт об увиденном, при стимуляции правого полушария такой ответ невозможен, но человек в состоянии найти и показать предъявленный ему объект, правильно выбрав его среди нескольких альтернатив, или выказать свое отношение к изображаемой сцене. Вместе с тем различия в зрительном восприятии при стимуляции разных полушарий, различия в зрительно-когнитивных нарушениях у больных с односторонними поражениями мозга и результаты многочисленных психофизиологических исследований на здоровых людях – все свидетельствует о том, что левое и правое полушария перерабатывают зрительную информацию и опознают зрительные образы неодинаково (обзоры: Л.И. Леушина с соавт., 1982; А.А. Невская, 1985; А.А. Невская и Л.И. Леушина, 1990].
Зрительная система человека анатомически организована так, что изображения экспонируемые в правой половине поля зрения, проецируются с обоих глаз в зрительную кору левого полушария, а из левой половины поля зрения - в соответствующую область правого полушария. Далее оба полушария могут обмениваться информацией. Исключить этот обмен у здорового человека невозможно, но можно его затруднить, усложнив условия наблюдения, так чтобы человек не мог узнавать предъявляемые стимулы со 100% вероятностью. В этих условиях обычно фиксируется разница в точности ответов (или в их скорости), когда стимул предъявлен слева от точки фиксации или справа от нее. Эти отличия и считают обычно проявлениями различий в работе правого и левого полушарий. Действительно: обмен информацией между полушариями требует дополнительного времени и сопряжен с некоторой потерей информации, поэтому при её дефиците, когда человек вынужден принимать решение по минимуму информации, увеличивается вероятность, что это решение принято в том полушарии куда прямо был адресован стимул. Отсюда следует, что при стимуляции левого поля зрения (ЛПЗ) более активно проявляет себя правое полушарие, а при стимуляции правого поля зрения (ППЗ) – левое полушарие.
В наших экспериментах, проведённых на здоровых взрослых испытуемых (все – правши), мы предъявляли изображение на короткое время (не более 100-140 мс), и вслед за стимулом следовал маскирующий стимул, чтобы оборвать процесс переработки зрительной информации, переключив зрительную систему на новую задачу. Вместе с тем, мы старались, чтобы условия опыта не создавали трудностей ни для памяти, ни для выбора и организации ответной реакции. Изображения предъявлялись по одиночке. Набор возможных стимулов был обычно известен наблюдателю. Использовался обычно более естественный для человека вербальный ответ: наблюдатель называл предъявленное изображение или записывал на специальном бланке его условное обозначение, но при этом не требовалось отвечать как можно быстрее и в случае сверки испытуемый мог исправить свой ответ.
Более подробно описание всех экспериментов и полученных результатов можно найти в книге А.А. Невской и Л.И. Леушиной (1990).
Для выявления межполушарных различий в переработке зрительной информации были поставлены следующие вопросы:
1) Одинаковы ли признаки, используемые в правом и левом полушариях при описании изображений?
2) Есть ли межполушарные различия в инвариантном опознании зрительных образов?
3) Есть ли различия в пространственно-частотном или гармоническом описании изображений в правом и левом полушариях?
На основании экспериментальных данных, полученных при исследовании этих проблем, предлагаются некоторые гипотезы относительно того, как организован процесс обработки зрительной информации и опознание зрительных образов в разных полушариях.
в левом и правом полушариях.
Если два полушария, описывая одни и те же изображения, выделяют в них неодинаковые признаки, то ошибки смешивания изображений могут быть неодинаковы, когда стимулы экспонируются в ЛПЗ и ППЗ. Были проанализированы систематические ошибки смешивания фигур в условиях кратковременного их предъявления в ЛПЗ и ППЗ.
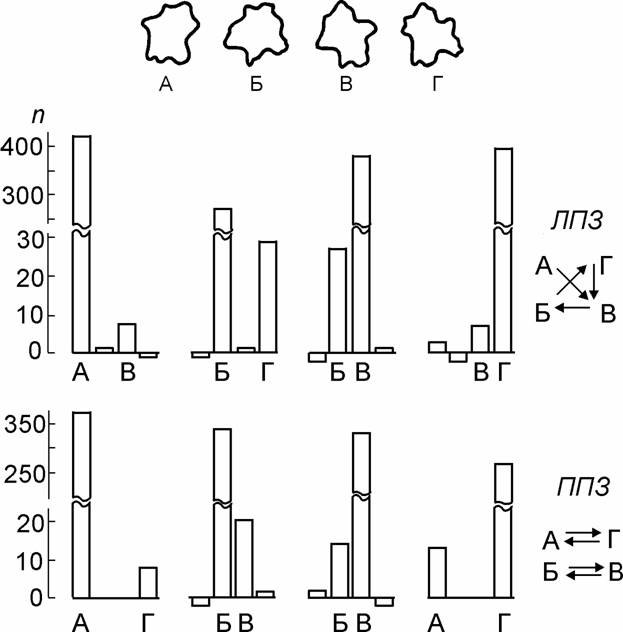
Наблюдатель выучивал небольшой набор изображений, например – 4 бессмысленных криволинейных фигуры, такие, как показаны на рис. 1.
Рис. 1. Распределение неслучайных (обусловленных стимулом) ответов о форме изображений, предъявленных в ЛПЗ и ППЗ.
Вверху – стимулы и их условные обозначения; n – число неслучайных ответов разных классов (А, Б, В, Г) о каждом из стимулов (отрицательные значения обусловлены статистической погрешностью при разложении экспериментальных матриц). Справа - схемы систематических ошибок для ЛПЗ и ППЗ (стрелки указывают, с каким изображением смешивается стимул. Испытуемый АН.
Эти фигуры предъявлялись многократно (от 800 и до 3000 проб у разных наблюдателей) в случайном порядке и с равными вероятностями в ЛПЗ и ППЗ. Длительности предъявления подбирали индивидуально для каждого испытуемого в диапазоне от времени, когда опознание было почти безошибочным до времени при котором вероятность правильных ответов снижалась почти до случайного уровня. Поскольку ответы «не знаю» были запрещены, наблюдатель был вынужден хотя бы иногда отвечать наугад. Поэтому, полученную в экспериментах матрицу ответов можно рассматривать как сумму двух матриц: матрицы случайных ответов (среди них могли быть и ответы случайно оказавшиеся правильными), и матрицы ответов, обусловленных стимулом. Метод разделения экспериментальной матрицы на две составляющих и техника расчётов, описаны в наших работах (Л.И. Леушина, А.А. Невская, 1978; 1990). Рассчитывали по всем клеткам матрицы максимально возможное число случайных ответов, но так чтобы это число нигде не превышало числа ответов данной категории, полученных в эксперименте (с точностью до некоторой погрешности, не превышающей по всей матрице 95% доверительного уровня). Вычитая из экспериментальной матрицы максимально возможную матрицу случайных ответов, получили матрицу ответов, обусловленных предъявленным стимулом.
Пример распределения только этих неслучайных ответов, приведен на рис. 1. для предъявления изображений в ЛПЗ и ППЗ. Как видно, среди неслучайных ответов значительную часть составляют правильные ответы, но присутствуют также и некоторые типы ошибок, число которых значительно превышает вероятность случайного их появления. Эти систематические ошибки возникают, очевидно, в случаях когда наблюдатель получил некоторую информацию о стимуле, но её оказалось недостаточно для выбора правильного ответа и человек называет близкое, похожее изображение. Эти систематические ошибки оказались различны при предъявлении изображений в ЛПЗ и ППЗ.
В таблице 1 суммированы данные о систематических ошибках у 11 наблюдателей, работавших с набором из 4 фигур, показанных на рис. 1 или с зеркальными вариантами этих же фигур.
Таблица 1
Систематические ошибки смешивания стимулов
|
ИСПЫТУМЫЕ
|
АЛФАВИТ ИЗОБРАЖЕНИЙ |
СИСТЕМАТИЧЕСКИЕ ОШИБКИ |
|
|
Левое полушарие |
Правое полушарие |
||
|
М.П. |
4Ф ? 3Р 4Ф ? 1Р |
АÛ Г Б Û В АÛ Г Б Û В |
А®В®Б®Г®А А®В®Б®Г®А |
|
И.А. |
4Ф ? 4Р, норм+зерк. |
АÛ Г Б Û В |
В®Б®Г®А |
|
А.Н. |
4Ф ? 4Р 4Ф ? 1Р |
АÛ Г БÛ В АÛ Г Б Û В |
А®В®Б®Г®В А®Б®В ®Г®В |
|
В.Г. |
4Ф ? 4Р, норм+зерк. |
АÛ Г Б Û В |
Г®А®В®Б |
|
Е.П. |
4Ф ? 4Р, зеркальн. |
АÛ Г Б Û В |
А®Б®Г®А |
|
А.К. |
4Ф ? 4Р, зеркальн. |
АÛ Г Б Û В |
? |
|
В.Я. |
4Ф ? 4Р, зеркальн. |
АÛ Г Б Û В |
ВÛА®Б®Г |
|
К.Я. |
4Ф ? 4Р |
АÛ Г Б Û В |
А®Б®Г®В®А |
|
П.Ф. |
4Ф ? 4Р |
А®В®Б®А |
АÛ Г Б Û В |
|
Н.А. |
4Ф ? 4Р |
А®Б®Г®В |
АÛ Г Б Û В |
|
И.Ч. |
4Ф ? 1Р, зеркальн. |
А®Б®В®Г |
АÛ Г Б Û В |
Выявлено следующее: 1) При предъявлении стимулов в ППЗ, когда они адресованы левому полушарию, у большинства наблюдателей (у 8 из 11) систематические ошибки оказались взаимно симметричны (например, фигуру «Б» наблюдатели постоянно смешивали с «В» и обратно: фигуру «В» они смешивали с «Б») При экспозиции тех же стимулов в ЛПЗ ошибки были несимметричны (например, фигура «Б» часто принималась за «Г», но не было ошибок противоположного направления). 2). Такой характер ошибок был выявлен независимо от условий эксперимента (моно- или бинокулярное наблюдение, вербальный ответ или показ на ответной карте, предъявление одного стандартного размера или же каждая из форм могла предъявляться в 3 или 4 вариантах разных размеров). 3). У 3 наблюдателей из 11 был получен прямо противоположный результат: взаимно симметричные систематические ошибки наблюдались при работе их правого, но не левого полушария. Заметим, что, если наблюдатели этой группы участвовали в других сериях экспериментов по межполушарной асимметрии, они также демонстрировали результаты прямо противоположные данным большинства испытуемых. 4). Взаимно-симметричные ошибки, характерные для левого полушария большинства наблюдателей оказались идентичны у всех 11 испытуемых: смешивались попарно фигуры «А» и «Г», «Б» и «В». Для другого полушария систематические ошибки, выявляемые у разных испытуемых оказались неодинаковы.
Сходные результаты были получены при опознании набора простых геометрических фигур и при опознании текстур: в одном полушарии (чаще – левом) среди систематических ошибок преобладали взаимно-симметричные и одинаковые у разных лиц, в другом полушарии такой симметрии не наблюдалось, и ошибки были различны у разных лиц.
Наличие двух типов систематических ошибок – стандартных и взаимно-симметричных в одном полушарии, варьирующих и несимметричных в другом - показал, что системы описания изображений в левом и правом полушариях организованы по-разному. Наличие взаимно-симметричных ошибок, очевидно, свидетельствует о том, что пара часто смешиваемых друг с другом фигур воспринимаются как похожие, их описания близки. Эта близость может быть представлена как расстояние в пространстве описаний. Следовательно, в этом случае можно говорить о существовании метрического пространства описания изображений и это пространство организовано одинаково у разных наблюдателей. В случае несимметричных ошибок (характерных чаще для правого полушария) представления о близости описаний различных изображений и о метрическом пространстве их описаний неприложимы. Можно думать, что каждое изображение из выученного набора описывается само по себе, вне связи с описаниями других изображений. Несимметричные ошибки различны у разных лиц, они могут быть неодинаковы и у одного и того же наблюдателя при опознании формы изображений, варьирующих по размеру, местоположению или по ориентации. Это свидетельствует о том, что зрительная система правого полушария может давать много разных описаний одного и того же объекта.
Представляется целесообразным рассмотреть выявленные различия между полушариями с точки зрения теории опознающих систем. Среди специалистов, разрабатывающих технические опознающие устройства существуют два основных подхода: дискриминантный (или классификационный метод) и структурный (структурно-лингвинистический) (E. Hunt, 1978; K. Fu, 1977; В.Д. Глезер, 1966, 1985).
При дискриминантном методе каждое изображение описывается большим набором признаков и представляется как точка в многомерном пространстве описаний. Предполагается, что все изображения, принадлежащие одному объекту, получают сходные описания и образуют компактную группу близлежащих точек в пространстве описаний (это условие выполняется, если признаки, составляющие оси этого пространства инвариантны к местоположению изображения, масштабным и другим преобразованиям). Задача обучения в этом случае состоит в установлении границы (гиперповерхности), разделяющей разные классы изображений - разные зрительные образы. Таков, например, принцип работы перцептрона – первой модели обучаемого распознающего устройства.
При структурном подходе изображение членится на составляющие его элементы и определяется как эти элементы расположены друг относительно друга. Задача обучения – определение правил, какие элементы могут или должны обязательно присутствовать во всех изображениях определенного зрительного образа, и по каким правилам эти элементы сочетаются один с другим. Для всякой задачи по распознаванию формируется специализированный «язык» описания изображений. Обозначения элементов можно уподобить словарю, а правила их сочленения – грамматике. Как в языке одно и то же явление может быть описано с использованием разных слов и разных грамматических конструкций, так и структурное описание как отдельных изображений, так и целых классов зрительных образов оказывается многообразным и гибким.
Возникло предположение, что мозг использует одновременно оба метода опознания зрительных образов, но они реализуются в разных полушариях: дискриминантный метод – преимущественно в левом полушарии, структурный - в правом.
Для экспериментальной проверки этого предположения мы постарались определить, что запоминают разные полушария, когда человек выучивает какой-то набор зрительных образов. Если опознание происходит по структурному методу, человек должен запоминать структуры тех образцов, которые давались ему при обучении, чтобы на основе конкретных описаний выработать общее правило построения зрительных образов. Если используется дискриминантный метод, не нужно помнить конкретные образцы, достаточно хранить в памяти сведения о положении границы, разделяющей зрительные образы друг от друга. Для экспериментов были использованы бессмысленные контурные фигуры и на их основе были построены ряды с постепенными переходами от одной фигуры к другой. Предварительная проверка показала, что эти ряды воспринимаются действительно как упорядоченные с плавными и достаточно равномерными переходами.
Испытуемые выучивали только 4 эталонные фигуры, а затем при краткой экспозиции им предъявляли в случайном порядке все стимулы трёх переходных рядов. Поскольку человек не знал о возможности предъявления измененных фигур, он относил каждый стимул к одному из известных ему эталонов.
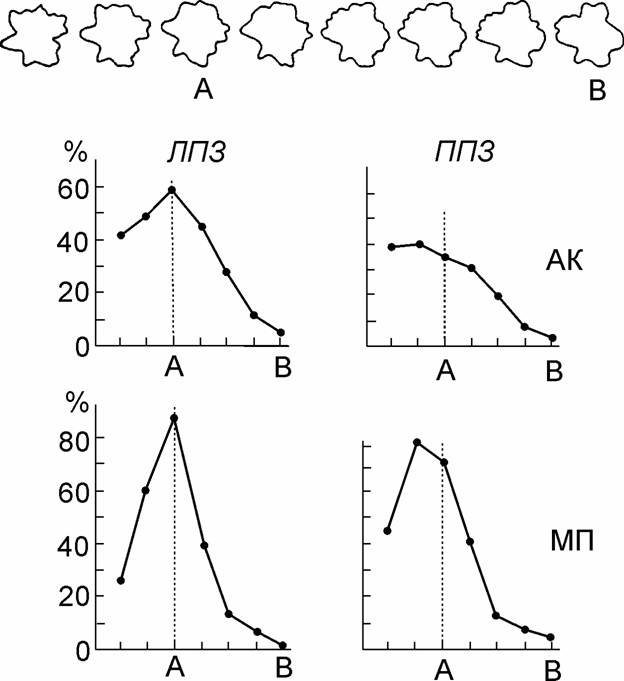
Рис. 2. Классификация фигур переходного ряда при выучивании эталонов А и В (усредненные данные для трех переходных рядов) в ЛПЗ и ППЗ.
Вверху – образец переходного ряда и положение в нем эталонных (выученных) стимулов. На графиках: по оси абсцисс – положение стимулов в переходном ряду, по оси ординат – процент ответов категории А. Испытуемые АК и МП.
На рис. 2 показан образец одного из переходных рядов, положение в нем выученных эталонов «А» и «Б» и результаты классификации всех переходных фигур при предъявлении их в ЛПЗ И ППЗ для двух наблюдателей. Для ЛПЗ, т.е. для правого полушария, максимальный процент правильных ответов категории «А» пришелся точно на эталонное изображение, любое отклонение стимула от выученного эталона ухудшало опознание. Но при экспозиции тех же изображений в ППЗ максимум правильных ответов категории «А» оказался смещён на 1-2 ступени в направлении от границы с другими эталонами, т.е. измененные фигуры узнавались левым полушарием даже лучше, чем выученные эталоны. Противоположный результат был зарегистрирован у наблюдателя П.Ф., который и в предыдущей серии экспериментов показал инверсную специализацию полушарий в обработке зрительной информации.
Таким образом, два полушария используют неодинаковые признаки при опознании. Правое полушарие хранит в памяти характеристики конкретных изображений, вырабатывая свой специфический «язык» для их описания. В левом полушарии формируется единое метрическое пространство описания изображений (одинаковое у разных лиц), и запоминаются лишь границы между областями этого пространства, которые соответствуют разным классам зрительных образов.
Очень похожие результаты были получены в исследовании Марсолек (С.K. Marsolec, 1995). В его экспериментах испытуемые заучивали 8 классов бессмысленных фигур, составленных соединениями или пересечениями нескольких прямолинейных отрезков; при обучении каждый класс был представлен 4 образцами, немного отличающимися друг от друга. Затем испытуемым предлагали классифицировать заученные фигуры, ранее не предъявлявшиеся прототипы этих фигур и новые, тоже ранее не предъявлявшиеся отклонения от тех же прототипов. Прототипы и новые варианты классифицировались более эффективно, если они предъявлялись в ППЗ, т.е. левому полушарию. Автор пришел к выводу, что в правом полушарии запоминаются конкретные изображения со всеми их деталями, но в левом полушарии функционирует относительно независимая система, которая формирует и хранит в памяти абстрактные зрительные образы.
Особенности инвариантного описания изображения
в левом и правом полушариях.
Существуют разные точки зрения относительно физиологических механизмов, обеспечивающих возможность инвариантного опознания видимых предметов при изменении положения их изображений на сетчатке, размера, ориентации, яркости. Более традиционным является представление, что зрительная система описывает объект конкретно, со всеми его особенностями, и лишь затем в процессе обучения на более высоких уровнях мозга формируется обобщение разных вариантов одного объекта (D.O. Hebb, 1949). Предполагается, что инвариантность обеспечивается мозговыми механизмами более высокого уровня, чем зрительная система. Согласно другой точке зрения в самой зрительной системе заложены механизмы, позволяющие описывать форму (а для большинства предметов именно форма является наиболее существенным и наиболее постоянным из воспринимаемых зрением свойств) инвариантно к местоположения, размеру, яркости и другим преобразованиям на сетчатке. Зрительный образ, с этой точки зрения, представляет собой обобщенное, лишённое конкретности описание формы видимого предмета. Но одновременно в зрительной системе существуют параллельные каналы, по которым передаются сведения о положении, размере и других особенностях видимого объекта (В.Д. Глезер, 1966; Л.И. Леушина, 1978; В.Д. Глезер с соавт., 1992). Многочисленные экспериментальные исследования свидетельствуют о том, что в зрительной системе реализуются оба рода описаний видимых объектов: абстрагированное, инвариантное к ряду преобразований описание формы объекта с параллельным независимым описанием других свойств изображения и единое полное конкретное описание изображения, в котором информация обо всех его свойствах неразделима. Задачей данного раздела исследований было выяснить, существуют ли межполушарные различия в инвариантности на уровне зрительного описания изображений, и как представлены два рода описаний (абстрагированное, инвариантное описание формы с наличием параллельных каналов для оценки других свойств и единое полное описание) – в разных полушариях.
Один из методов экспериментального исследования межполушарных различий в инвариантности состоял в следующем: наблюдателям предъявляли стимулы, варьирующие по двум параметрам, например, по форме и положению в поле зрения или по форме и размеру, и требовалось одновременное опознание обоих свойств предъявленного стимула. Мы исходили из следующей гипотезы: если в зрительной системе действуют механизмы инвариантного описания формы и есть раздельные каналы для обработки информации о форме, о местоположении стимула, о его размере и т.д., то оценка всех этих свойств производится независимо друг от друга, и тогда вероятность правильного опознания одного из этих свойств не должна зависеть от того правильно или неправильно опознано другое свойство. В случае единого полного описания изображения все его свойства взаимосвязаны, и тогда вероятность правильного опознания любого из них должна существенно зависеть от того, верно или неверно оценено другое свойство.
Для примера в таблице 2 представлены результаты одного эксперимента.
Таблица 2
Опознание формы фигур в ЛПЗ и ППЗ в случаях неверно
оцененного местоположения стимулов.
Испытуемая Н.М. Опыты 1- 6 (Изображения предъявлены по 780 раз в каждое поле зрения.
За всю серию форма опознана правильно 605 раз в ЛПЗ и 565 раз в ППЗ. Местоположение оценено неверно 194 раза в ЛПЗ и 314 раз в ППЗ).
ЛПЗ c 2 = 0,06 n=1; Р>0,8
Ответ
Стимул |
квадрат |
звезда |
Всего
стимулов |
|
Квадрат |
39 |
24 |
63 |
|
Звезда |
77 |
54 |
131 |
|
Всего ответов |
116 |
78 |
194 |
ППЗ c 2 = 11,73 n=1;Р<0,001
|
Ответ
Стимул |
квадрат |
звезда |
Всего
стимулов |
|
Квадрат |
111 |
59 |
170 |
|
Звезда |
63 |
81 |
144 |
|
Всего ответов |
174 |
140 |
314 |
Набор стимулов состоял из 2 фигур размером около 1о, и каждая из них могла появится в одном из четырёх мест поля зрения в 3о от точки фиксации. Длительность предъявления подбирали так, чтобы обеспечить уровень правильных ответов около 60-70%. Анализировали ответы о форме изображения только в случаях неверной оценки его местоположения. Казалось бы - если человек грубо ошибся в месте предъявления стимула (например, фигура была предъявлена справа вверху, а человек ответил, что она была слева внизу), - как он может узнать форму фигуры? Однако проверка по методу c- квадрат показала, что для ЛПЗ ответы о форме, действительно, случайны, но для ППЗ предположение о случайности ответов о форме отвергается с высокой степенью достоверности. Таким образом, когда стимул адресован левому полушарию форма его может быть опознана даже в случаях неверной оценки его местоположения, в другом поле – только в случаях правильной его оценки. У большинства испытуемых (у 7 человек) инвариантное распознание формы осуществлялось в ППЗ, т.е. в левом полушарии, у двоих – в ЛПЗ, т.е. в правом полушарии, у одного наблюдателя инвариантное опознание было возможно при предъявлении стимулов в обоих полях зрения. Отметим, что условия наблюдения (число и сложность опознаваемых форм, число возможных местоположений стимулов, моно- или бинокулярное наблюдение) не отразились на результатах.
Другой пример: набор стимулов состоял из 4 форм (бессмысленные криволинейные фигуры), и каждая из них могла быть предъявлена в одном из четырёх размеров – в диапазоне от 4о до 8о . Изображения предъявляли по одному в случайном порядке и с равной вероятностью в ЛПЗ и в ППЗ. Наблюдатель должен был опознавать и форму и размер изображений. Чтобы исключить влияние несимметричного положения контуров фигуры в ЛПЗ и ППЗ по отношению к точке фиксации, чередовали серии с предъявлением фигур одной ориентации и их зеркальных вариантов, повернутых вокруг вертикальной оси, результаты этих серий суммировали. Длительности предъявления стимулов подбирали индивидуально так, чтобы для каждого наблюдателя иметь 5-7 разных длительностей в диапазоне от почти безошибочного опознания до уровня случайного гадания.
Результаты одного наблюдателя (В.Г.) представлены на рис. 3.
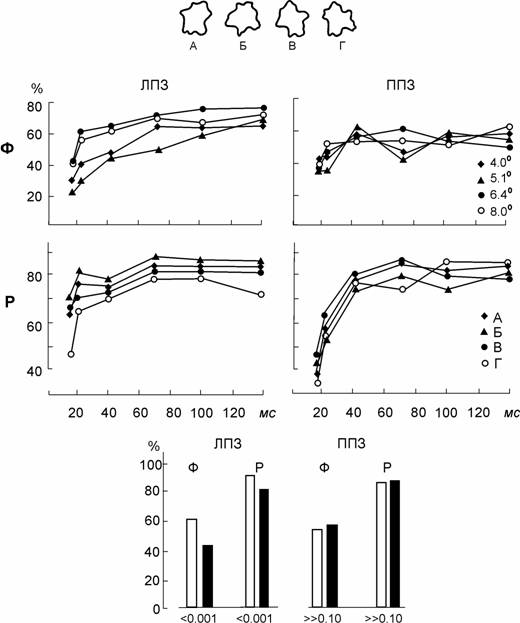
Рис. 3. Одновременное опознание формы изображений (Ф) и их размера (Р) в ЛПЗ и ППЗ.
Вверху – примеры изображений и их условные обозначения. На графиках – временные характеристики опознания формы для разных размеров изображений (4° - 8°) и опознания размера для изображений разных форм (А – Г). По оси абсцисс – длительность предъявления стимулов, по оси ординат – процент правильных ответов. Столбики внизу – усредненные по всем длительностям предъявления вероятности правильного опознания формы (Ф) и размера (Р) в случаях правильно опознанного второго свойства (светлые столбики) и в случаях ошибок в опознании второго свойства (темные столбики). Цифры под столбиками – вероятность отсутствия различий между этими случаями.
Видно, что размер изображений, предъявляемых в ППЗ (т.е. левому полушарию) не оказывал никакого влияния на опознание формы: кривые вероятности опознания формы для всех размеров идут вместе. Но при тех же условиях для ЛПЗ (т.е. для правого полушария) выявляется зависимость опознания формы от размера: кривые опознания формы при разных размерах упорядочены, и можно оценить, при каком размере изображения его форма опознаётся лучше, при каком - хуже. Такая же закономерность выявлена и для оценки размера изображений разных форм: в ППЗ никакие вариации формы не оказывают влияния на оценку размера, но в ЛПЗ вероятности оценки размера оказываются неодинаковы для разных фигур. Если сравнить вероятности правильной оценки любого из этих свойств в случаях, когда другое свойство оценено правильно или ошибочно, то для ППЗ (для левого полушария) не выявляется никакого влияния опознания одного свойства на другое, но в ЛПЗ (в правом полушарии) существует положительная корреляция: форма опознаётся лучше, когда размер оценен правильно, а размер оценивается лучше при правильно опознанной форме. Можно сделать заключение, что для левого полушария данного наблюдателя характерны инвариантное к масштабным преобразованиям описание формы при наличии раздельных каналов для опознания формы и размера, но в правом его полушарии осуществляется полное конкретное описание изображения без разделения на каналы.
Такой результат получился у 10 из 13 обследованных наблюдателей: у 3 человек была отмечена инверсная специализация полушарий (инвариантное описание формы и независимые каналы в правом полушарии, единое полное описание изображений – в левом) .
Различия между двумя полушариями в инвариантности четко выявляются в первые опытные дни. При продолжительном исследовании, особенно если от испытуемых требуется ответ только о форме изображения, игнорируя вариации его размера, картина может меняться: форма изображения постепенно начинает опознаваться независимо от размера в обоих полях зрения. Следовательно, инвариантное по отношению к размеру описание формы может сформироваться вторично под влиянием тренировки в исходно неинвариантном полушарии. Скорость формирования вторичной инвариантности очень различна у разных лиц: иногда она начинает проявляться уже после 3-5 дней исследования, но у некоторых испытуемых зависимость опознания формы от размера изображения в одном поле зрения сохранялась ещё и после двадцати опытных дней. Вторичная инвариантность может быть разрушена, если потребовать от испытуемого отвечать не только о форме, но и о размере предъявленного изображения.
Формирование вторичной инвариантности в исходно неинвариантном полушарии мы наблюдали также у некоторых (но не у всех) испытуемых, опознававших форму изображения при вариациях его местоположения (от испытуемых требовался отчёт об обоих этих свойствах стимула). Так наблюдатель Н.М. (результаты первых 6 опытах с ней приведены в табл. 2) постепенно научилась узнавать форму изображений в случаях ошибочной оценки их местоположения не только в ППЗ, но и в ЛПЗ. Для этого ей потребовалось около 4000 предъявлений. После длительной тренировки инвариантное, независимое от местоположения опознание формы (а также и оценка местоположения стимула независимо от его формы) в ЛПЗ, т.е. в правом полушарии, стали даже лучше, чем в исходно инвариантном левом полушарии (рис. 4). Но тренировка мало повлияла на успешность работы левого полушария.
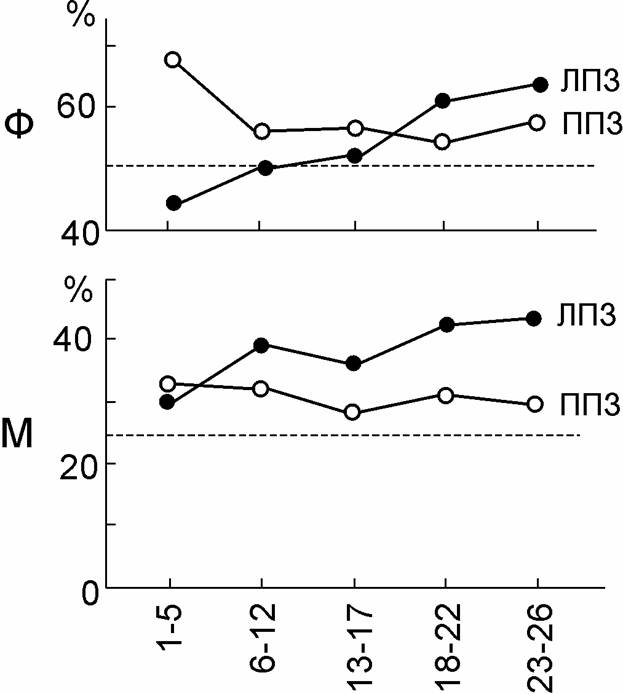
Рис. 4. Тренировка опознания формы (Ф) и местоположения (М) геометрических фигур в ЛПЗ и ППЗ.
Ф – опознание формы фигур в случаях неверно оцененного их местоположения (каждая точка – результат 200-350 проб); М – опознание местоположения фигуры в случаях неверно опознанной ее формы (каждая точка – результат 100-250 проб). По оси абсцисс – дни тренировки; по оси ординат – процент правильных ответов. Штриховые линии, параллельные оси абсцисс – уровень случайного гадания. Испытуемый НМ.
Формирование при обучении и тренировке вторичной инвариантности удалось хорошо проследить в серии экспериментов по опознанию формы фигур при изменении их ориентации.
В этих экспериментах наблюдатели не были знакомы с набором предъявляемых изображений. Незнакомая бессмысленная криволинейная фигура появлялась на короткое время в ЛПЗ или ППЗ, сопровождаясь маскирующим стимулом. Спустя 0.5 с на экране появлялась ответная карта с четырьмя фигурами на ней. Одна из них по форме точно соответствовала тестовой фигуре, но могла отличаться от неё по ориентации – поворотом на любой угол от 0° до 180° как по, так и против часовой стрелки. Три другие фигуры на ответной карте отличались от тестовой фигуры по форме, но имели одинаковый с ней какой-то фрагмент контура. Задача наблюдателя была найти на ответной карте фигуру идентичную по форме тестовой, и записать её номер; ответа об ориентации фигур не требовалось. Принимались меры, чтобы, чтобы наблюдатели не могли выучить тестовые фигуры или последовательность их предъявления. Для этого использовался достаточно большой набор стимулов – 16 тестовых фигур разной формы, и в течении дневного опыта каждая из них предъявлялась в 8 различных ориентациях, повторяясь только по одному разу в ЛПЗ и ППЗ. Таким образом за один опыт в каждом поле зрения демонстрировались по 128 неповторяющихся стимулов; ответные карты также постоянно менялись.
Исследование было проведено на 32 испытуемых, все взрослые здоровые люди с нормальным или коррегированным до нормы зрением, все правши. 24 человека были «наивными» наблюдателями, никогда ранее не участвовавшими в подобных экспериментах и не знавшие их цели. 8 человек, составляющих контрольную группу, участвовали ранее в различных экспериментах по межполушарной асимметрии при зрительном опознании, и для каждого из этих наблюдателей было известно, какое его полушарие обладает исходной инвариантностью к местоположению стимула или масштабным его преобразованиям.
Четкие различия по вероятности правильного опознания формы повернутых фигур при экспозиции их в ЛПЗ и ППЗ были выявлены у 29 наблюдателей из 32. Большинство – 23 человека, в том числе все 7 наблюдателей из контрольной группы, у которых первично инвариантным по данным предшествующих исследований было их левое полушарие, давали более высокий процент правильного опознания формы, когда стимулы предъявлялись в ППЗ, хотя в первый опытный день этот процент ни у кого не превышал 50%. При предъявлении тех же стимулов в ЛПЗ процент правильных ответов о форме повёрнутых фигур в первый опытный день у 18 испытуемых не отличался значимо от вероятности случайного угадывания (25%) .
Таким образом, у большинства наблюдателей инвариантное опознание формы фигур при изменениях их ориентации обеспечивается в первые опытные дни работою механизмов левого полушария. Правое полушарие в это время или совсем не узнаёт формы повернутых фигур, или делает это весьма плохо. Противоположные результаты мы получили у 6 лиц, включая одного наблюдателя из контрольной группы с первично инвариантным правым полушарием.
Тренировка – а с каждым испытуемым опыты повторялись от 8 до 21 раза – оказывала неодинаковое влияние на опознание повернутых фигур, адресованных в разные полушария.
У всех наблюдателей с первично инвариантным левым полушарием тренировка сравнительно мало влияла на работу этого полушария, но от опыта к опыту увеличивался процент правильных ответов для стимулов в ЛПЗ, т.е. существенно возрастали возможности правого полушария узнавать форму повернутых фигур, и постепенно именно правое полушарие стало ведущим, преобладающим в решении данной задачи (рис. 5, а, в, г).
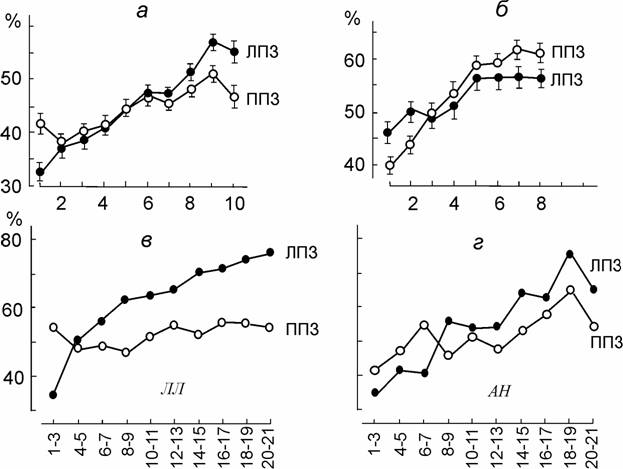
Рис. 5. Тренировка опознания повернутых изображений, предъявляемых в ЛПЗ и ППЗ; суммарные данные для всех углов поворотов фигур.
а – усредненные результаты двенадцати «наивных» испытуемых с левым первично инвариантным полушарием; б - усредненные результаты трех испытуемых с правым первично инвариантным полушарием; в, г – индивидуальные результаты длительной тренировки двух испытуемых из контрольной группы (ЛЛ и АН) с левым первично инвариантным полушарием. По оси абсцисс – дни тренировки; по оси ординат – процент правильных ответов.
У лиц с инверсной специализацией полушарий тренировка, напротив, резко улучшала опознание повернутых изображений в ППЗ, и преимущество в опознании переходило от правого полушария к левому (рис. 5, б) Скорость выработки вторичной инвариантности сильно различалась у разных испытуемых. Можно думать, что у некоторых особенно быстро тренирующихся наблюдателей мы просто не успевали замечать перехода преимущества от одного полушария к другому и из-за этого результаты их трактовались как неясные, неопределенные.
Итак, инвариантное к изменениям ориентации опознание изображений возможно в обоих полушариях, но в первично инвариантном полушарии для этого существуют готовые механизмы, а в исходно неинвариантном полушарии такие механизмы начинают работать лишь после обучения и тренировки. И эти вновь сформировавшиеся механизмы обеспечивают опознание повернутых фигур на более высоком уровне, чем механизмы первичной инвариантности. Однако, встаёт вопрос: как может происходить обучение в условиях когда наблюдатель не получает информации относительно того правилен или ошибочен был его ответ о форме повернутого изображения? Можно думать, что в роли учителя здесь выступает первично инвариантное полушарие, где есть, хотя и несовершенные, но всё же готовые механизмы инвариантности.
Подводя итог всем исследованиям инвариантного опознания в левом и правом полушариях, мы можем отметить следующее. Обнаружены два типа инвариантности, связанные с разными полушариями. Исходная или первичная инвариантность мало зависит от обучения и тренировки. Можно думать, что механизмы, её обеспечивающие, являются врождёнными, генетически заданными. Вторичная инвариантность возникает только после обучения наблюдателя решению определённой задачи. Наличие разных типов инвариантности установлено не только у человека, но и у животных. На кошках при удалении левого или правого полушария было показано, что инвариантность к изменениям фигур по размеру и ориентации существует исходно в их левом полушарии, а в правом полушарии она может вырабатываться при обучении (Н.В. Праздникова с соавт., 1997).
У большинства людей (75 – 80%) первичная инвариантность тоже связана с работой левого полушария, а вторичная – правого. Однако, примерно у 20% наблюдателей отмечена инверсия в специализации полушарий, эта инверсия не связана с моторным левшеством, - все наши испытуемые были правшами. Характер специализации полушарий не изменялся со временем, он оказался одинаков при опознании изображений, варьирующих по местоположению, размеру или ориентации. Можно отметить также жесткую корреляцию между типом инвариантности и характером ошибок смешивания фигур: для полушария с первичной инвариантностью были типичны взаимно-симметричные стандартные ошибки, и, по нашим предположениям, именно это полушарие использует дискриминантный метод опознания зрительных образов.
Исходно инвариантное описание изображений существует, по-видимому, только для немногих, наиболее простых, но и наиболее часто встречающихся преобразований как вариации по местоположению и размеру. Изменения ориентации изображений - повороты более чем на 30° уже сильно снижают вероятность правильного опознания их формы с помощью механизмов первичной инвариантности. Вторичная инвариантность, по-видимому, может быть сформирована и для более сложных преобразований. Например, на больных с расщеплённым мозгом показано, что в левом полушарии обобщаются только варианты изображений с линейными преобразованиями, в то время как в правом полушарии возможны обобщения при линейных, аффинных, проективных и топологических преобразованиях (L. Franco, R.W. Sperry, 1977). Возможно, что превосходство правого полушария в опознании индивидуальных лиц или букв рукописного шрифта (M.P. Bryden, F. Allard, 1976) связано с тем, что форма этих объектов не является жесткой, т.е. мы имеем дело с более сложными формами вариабельности. Некоторые клиницисты отмечали при зрительных агнозиях различия в опознании больными живых и неживых объектов, причем узнавание первых сильнее нарушалось при дефектах правого полушария (H. Hecaen, 1969).
Гипотезы о нейрофизиологических механизмах, обеспечивающих врожденную инвариантность, предложены В.Д. Глезером (В.Д. Глезер, 1993, V.D. Glezer, 1995). Они основаны на пространственно-частотном описании изображений и модульной организации зрительной коры Мы рассмотрим их в следующем разделе. Формирование вторичной инвариантности можно объяснить на основе представлений Хэбба (D.O. Hebb, 1949): при показе различных вариантов одного объекта первоначально все они описываются по-разному, но по мере установления ассоциаций и формирования нейронных ансамблей всё более высокого уровня из полных описаний изображений исключаются те характеристики, вариации которых оказываются несущественны для данной задачи, и остаётся укороченное описание общее для целого класса изображений.
Межполушарные различия в гармоническом
пространственно-частотном описании изображений. Нейрофизиологические основы межполушарной
асимметрии в зрительных процессах.
Чтобы понять, что лежит в основе межполушарных различий в процессах переработки зрительной информации, желательно было бы проследить все этапы этой переработки, начиная с того уровня, где впервые появляются различия между полушариями.
В зрительной коре происходит переход от поточечного описания видимого мира к распределенному описанию: каждый корковый нейрон описывает распределение освещённости в некоторой области видимого пространства – в своём рецептивном поле. Согласно модели В.Д. Глезера (1993, 2000) рецептивные поля нейронов стриарной коры представляют собой двумерные решетчатые фильтры пространственных частот, осуществляющие кусочное Фурье-разложение изображения. Набор нейронов с рецептивными полями одного размера, направленный на один и тот же участок поля зрения, составляет модуль. Поскольку в модуль входят нейроны, реагирующие на разные ориентации и частоты, такой модуль даёт локальное спектральное описание своего участка зрительного поля.
Модули престриарной коры организованы по тому же принципу, и тоже выдают спектральное описание определённого участка поля зрения, но если стриарные модули описывают распределение освещённости, то престриарные модули описывают распределение возбуждений в нейронах предыдущего уровня.
Модули различной величины перекрывают один и тот же участок поля зрения, и благодаря механизмам внутри- и межмодульного торможения наиболее активным оказывается тот из них, чьё положение, размеры и конфигурация в максимальной степени соответствуют относительно однородному участку в поле зрения. Таким образом, начинается сегментация зрительного поля на участки, соответствующие отдельным изображениям, образам или подобразам.
Для описания каждого объекта выбирается свой модуль, наиболее подходящий по размеру и местоположению, но на подбор модуля, по-видимому, оказывают влияния и ожидания наблюдателя. Так, если человек ожидает появления стимула даже очень малого размера где-то в обширной области пространства, он может активизировать модули с большими рецептивными полями, чтобы иметь возможность анализировать всю область возможного появления стимула. Но если место появления стимула известно заранее, можно воспользоваться модулем малого размера.
Характеристики линейных нейронов, составляющих модуль, оказываются строго упорядоченными (В.Д. Глезер, В.Е. Гаузельман, 2001) и на выходе модуля выделяются 6 ориентаций, 5 гармоник и 2 фазы (четная и нечетная гармоники). Таким образом, каждое изображение или же его деталь, выделенные данным модулем описываются по 60 признакам, т.е. представляются как точка в 60-мерном пространстве. Такой способ представления информации позволяет перейти к инвариантному описанию формы изображения. Все изображения одинаковые по форме, но различающиеся местоположением на сетчатке или размером, описываются разными модулями, но на выходе любого из них получается одно и то же описание гармонического состава изображения. Иными словами – выходные сигналы модуля представляют собой признаки, инвариантные к масштабу и параллельному переносу. В то же время, зная какой именно модуль возбужден наиболее сильно при предъявлении данного конкретного изображения, нетрудно определить его местоположение и размер, что создаёт предпосылки для разделения каналов описания формы, местоположения, размера и других параметров изображения.
Существование межполушарных различий на уровне стриарной коры кажется нам маловероятным: стриарные области двух полушарий не связаны прямо комиссуральными связями, и если бы какая-то информация не воспринималась одним из полушарий, она оказалась бы безвозвратно потеряна. В этом случае мы видели бы одни и те же объекты по-разному в ЛПЗ и ППЗ. Например, если бы полушария различались по пространственно-контрастной чувствительности, это должно было бы проявиться в различиях по остроте зрения между левым и правым полуполями, но таких данных нет.
Выделение объекта из фона (кроме, может быть, некоторых наиболее сложных случаев) осуществляется по-видимому, стриарной корой, и в этом случае межполушарные различия не выявляются (В.Д. Глезер с соавт., 2002).
Что касается престриарной коры, то на этом уровне путём подбора модуля подходящего размера и конфигурации, очевидно, уже осуществлено выделение некоего объекта для дальнейшего анализа – «что мы попытаемся опознать». На этом уровне возможно появление межполушарных различий. Москович (M. Moscovich, 1979) анализируя самые различные данные о межполушарной асимметрии, так же приходит к заключению, что престриарная кора или же стык её с задне-височной корой является наиболее вероятным локусом для появления межполушарных различий.
Можно ожидать, что полушария будут различаться не в чувствительности к разным пространственным частотам, а в использовании разных областей пространственно-частотного описания изображений.
Возникло предположение, что левое полушарие использует в основном низкочастотную часть пространственно-частотного спектра изображений, а правое полушарие – его более высокочастотную часть. Для проверки этого предположения были проведены эксперименты. Наблюдателям предъявляли полутоновые фотографические изображения обычных предметов с отфильтрованными до разного уровня высокими пространственными частотами (рис. 6).
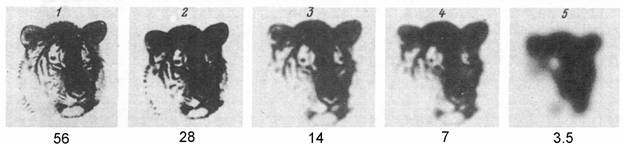
Рис. 6. Примеры тестовых изображений с разными уровнями отфильтровывания высоких пространственных частот.
1-5 – условные обозначения степени фильтрации изображений; цифры внизу – число сохранившихся гармоник в циклах на изображение.
Использовали наборы из 5 изображений и 5 уровней фильтрации, так что в изображении сохранялись гармоники 3.5, 7, 14, 28 или 56 циклов на изображение. При кратковременном предъявлении этих изображений с последующей маскировкой наблюдатель должен был ответить, где был предъявлен стимул - в ЛПЗ или ППЗ, назвать изображение и оценить условным номером по пятибальной шкале степень его размытости, т.е. уровень отфильтровывания высоких частот. Предлагая эти задания, мы полагали следующее: чтобы оценить степень размытости изображений (уровень фильтрации) необходимо определить, какие самые высокие пространственные частоты или гармоники ещё присутствуют в изображении. Для обнаружения стимула достаточно самых низких пространственных частот или гармоник. Для опознания формы может использоваться широкая полоса частот, - хотя предметные изображения часто распознаётся по 3-4 гармоникам, было показано, что присутствие в них более высоких гармонических составляющих облегчает опознание.
В различных экспериментах изображения предъявлялись в двух размерах – 7° или 1°55¢. Хотя при этом абсолютные значения присутствующих в стимуле пространственных частот менялись в три с лишним раза, результаты обоих экспериментов оказались удивительно схожими. Существенны оказались не абсолютные значения пространственных частот (в циклах на градус), а число гармоник, сохранившихся в изображении ( в циклах на изображение).
При обнаружении стимула ошибки были редки и встречались лишь при самых коротких длительностях предъявления (10 - 15 мс); больше ошибок было отмечено при предъявлении стимулов в ППЗ (Рис. 7).
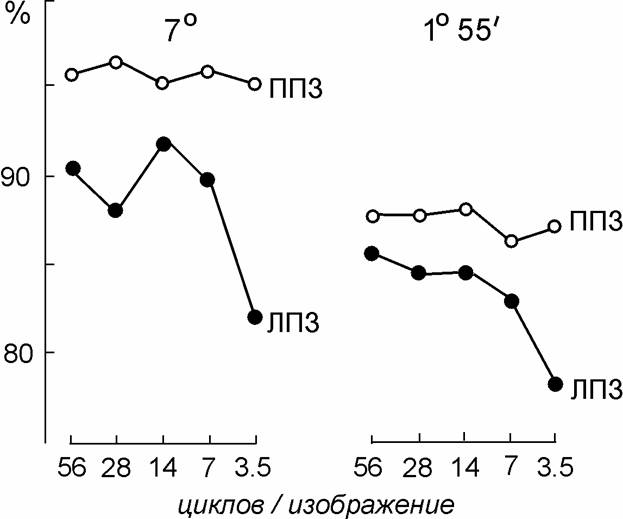
Рис. 7. Вероятность правильного обнаружения изображений с отфильтрованными высокими пространственными частотами в ЛПЗ и ППЗ.
По оси абсцисс – число гармоник, сохранившихся в изображении; по оси ординат – процент правильных ответов о местоположении стимула. 7° и 1°55¢ - размеры изображений; каждая точка – результат 300 проб при длительностях предъявления 10-30 мс. Испытуемый АН.
Хотя отфильтрованные (размытые) изображения субъективно воспринимались как более слабые, степень фильтрации совсем не влияла на их обнаружение в ППЗ, а при предъявлении стимулов в ЛПЗ вероятность их обнаружения лишь немного снижалась при самой сильной степени размытости (когда в стимуле оставалось всего 3,5 гармоники). Это показывает, что обнаружение стимула производится, действительно, на основе выделения низкочастотной части его спектра (особенно в ППЗ, т.е., в левом полушарии), и левое полушарие выделяет эти низкие гармоники лучше, чем правое.
Оценка степени размытости была более точной, когда стимулы предъявлялись в ЛПЗ, т.е. адресовались правому полушарию (Рис. 8), и только при различении двух наиболее размытых изображений – с границами 7 и 3, 5 цикла на изображение – небольшое преимущество имело левое полушарие.

Рис. 8. Оценка четкости – размытости изображений, предъявленных в ЛПЗ и ППЗ.
Зависимость среднего количества получаемой информации (I) о степени размытости изображений от длительности предъявления стимулов. 7° и 1°55¢ - размеры изображений; каждая точка – результат 800 или 550 проб. Испытуемый АН.
Таким образом, хотя в целом правое полушарие оценивало четкость – размытость точнее, в случаях, когда стимулы не содержали высоких гармоник, задача лучше решалась правым полушарием.
При опознании формы изображения межполушарных различий по проценту правильных ответов не было выявлено (хотя это и не служит доказательством того, что оба полушария используют при опознании одну и ту же полосу пространственных частот ).
Таким образом, эксперименты показали, что при описании изображений в правом полушарии используются высокие гармоники изображения, а в левом – низкие. Подчеркнём, что речь идёт именно о гармониках, но не об абсолютных значениях пространственных частот. А это означает, что межполушарные различия возникают на том уровне, когда уже выделен некий объект для анализа - изображение.
Анализируя полученные результаты, мы высказали следующее предположение (А.А. Невская, Л.И. Леушина, 1990, 1995): модули обоих полушарий организованы и работают единообразно, но отличаются размерами рецептивных полей, - в левом полушарии эти поля крупнее, чем в правом. Поскольку каждый модуль выделяет ограниченное число пространственных частот, то модули левого полушария с большими рецептивными полями описывают объект как единое целое, грубо, без мелких деталей, в то время как более мелкие модули правого полушария описывают тот же объект по частям, но зато описание получается более точным и подробным.
Из гипотезы о том, что межполушарные различия в зрительном восприятии базируются на различиях в размерах рецептивных полей корковых модулей, вытекают два предположения. Во-первых, можно ожидать, что маленькие изображения должны лучше опознаваться в правом полушарии, большие – в левом (мы не рассматриваем очень больших объектов, которые осматриваются только по частям с последовательным переводом взора с одной части на другую). Во-вторых, при очень кратких экспозициях стимула, когда возможности его восприятия ограничены малой энергией сигнала, преимущество должно иметь левое полушарие, - чем больше рецептивное поле, тем быстрее происходит накопление энергии, позволяющее выделить сигнал из шума, хотя при таком коротком времени ещё почти невозможны точное описание и опознание формы изображения. Оба эти следствия получили подтверждение в экспериментах (А.А.Невская, Л.И. Леушина, 1995) (рис. 9 и 10).

Рис. 9. Влияние размеров изображений (при одном и том же их наборе) на опознание их формы в ЛПЗ и ППЗ.
Вверху – примеры изображений. 2.8° и 6.0° - размеры изображений. На графиках - зависимость среднего количества получаемой информации (I) о форме изображения от длительности предъявления. Испытуемый ЕП.

Рис. 10. Влияние длительности экспозиции на вероятность опознания формы изображений в ЛПЗ и ППЗ.
На графиках – зависимость процента правильных ответов от длительности предъявления изображений. Данные трех испытуемых (ВГ, ИА, АН); n – число проб на одну точку.
В.Д. Глезером позднее была предложена другая версия относительно нейрофизиологических оснований, вызывающих межполушарные различия
(В.Д. Глезер, 2000). В зрительной коре кроме нейронов с линейными свойствами, которые могут осуществлять кусочное Фурье-разложение изображения, существуют также многочисленные нелинейные нейроны. Их можно считать специализированными детекторами, выделяющими линии определённой ориентации, полосы, углы, перекрестия, соединения контуров, пики, изгибы и т.д. Было введено понятие примитивов или локальных признаков – далее неразложимых элементов изображения. К числу примитивов относят также и текстуры. По мнению В.Д. Глезера, кроме примитивов, выделяемых врождёнными, генетически закреплёнными механизмами, и некоторые другие часто встречающиеся элементы изображений могут под влиянием тренировки приобрести свойства примитивов.
Механизмы пространственно-частотного анализа изображений и выделения примитивов сосуществуют в стриарной коре. Предполагается, что отсюда пространственно-частотные описания направляются в нижневисочную кору левого полушария, где конструируется многомерное пространство, оси которого являются инвариантными признаками. Область этого пространства представляет собой абстрактное инвариантное описание зрительного образа. Эта область выделяется нелинейными нейронами левой нижневисочной коры, которые определяют её границы, т.е. гиперповерхности, отделяющие область одного зрительного образа от других. В памяти сохраняются только значения этих границ. Но по ним невозможно восстановить первоначальное описание изображения. Описание по примитивам направляется из стриарной коры в правое полушарие, и правая нижневисочная кора с помощью пространственных операторов заднетеменной коры строит из примитивов полное структурное описание изображения со всеми его конкретными особенностями и деталями. Инвариантность здесь исходно отсутствует, но она может формироваться при обучении и тренировке путём выработки ассоциаций (согласно представлениям Хэбба). При изменении задачи классификации эта вторичная инвариантность может разрушиться, но на основе полных описаний изображений формируются новые обобщения, более адекватные для новой задачи.
Заметим, что обе вышерассмотренные концепции достаточно сходны: составляют ли основу межполушарных различий разные размеры рецептивных полей модулей правого и левого полушарий, или же различия коренятся в линейных – нелинейных свойствах нейронов стриарной коры, посылающих свои сигналы в разные полушария - следствия оказываются одинаковыми: Левое полушарие описывает видимый объект целиком, довольно грубо, но зато оно способно выделять инвариантные признаки формы. Правое полушарие даёт более подробное детализированное описание структуры этого же объекта, но инвариантность здесь отсутствует и может формироваться лишь вторично при обучении.
Зрительный образ в левом полушарии представлен коротким кодом, обозначающим только к какому из известных человеку классов - зрительных образов принадлежит данное изображение. Когда стоит задача найти знакомый объект среди множества незнакомых и опознать его, это быстрее и лучше делает левое полушарие. Но если требуется конкретизация, различение деталей и частных особенностей знакомого объекта, то превалирует правое полушарие (Н.Д. Чернова с соавт., 2000). Зрительные образы левого полушария благодаря своей краткости удобны для оперирования ими в мышлении. Высокий уровень абстрактности этих образов облегчает переход к абстракциям ещё более высокого уровня – к формированию понятий и обозначающих их слов. Разделение в левом полушарии каналов для инвариантного описания формы объекта и его пространственных характеристик (местоположение, размер, ориентация, движение) может составлять основу для предметных и предикатных номинаций в языке. Итак, можно думать, что характер зрительных образов, присущих левому полушарию (а также и образов в других сенсорных системах, где тоже выявлены межполушарные различия, во многом сходные с тем, что известно для зрения), создаёт предпосылки для формирования механизмов языка и речи именно в левом полушарии.
Зрительные образы правого полушария, сохраняющие сведения о структуре видимого объекта и о всех его частных особенностях, информационно богаче, чем левополушарные образы. Поэтому правое полушарие доминирует во всех задачах, где требуется мысленное оперирование с наглядным материалом: нарисовать увиденный и правильно опознанный объект, представить мысленно те или иные его трансформации, сформировать разнообразные обобщения на основе внешнего сходства или сходства структур.
Но в нормальных условиях у здорового человека оба полушария работают одновременно, постоянно обмениваясь информацией, дополняя возможности друг друга и помогая друг другу.
Глезер В.Д. Механизмы опознания зрительных образов. М., Л., 1966, c. 204
Глезер В.Д Зрение и мышление. Изд. 2-ое, перераб., 1993, 341 с.
Glezer V.D. (Глезер В.Д.) Vision and mind: modelling mental functions. Lawrens Erlbaum Ass. Publ., Mahwah N.Y., 1955, 274 p.
Глезер В.Д. О роли пространственно-частотного анализа, примитивов и межполушарной асимметрии в опознании зрительных образов. //Физиология человека, 2000, т. 26, №5, с. 145-150.
Глезер В.Д., Гаузельман В.Е. Модули стриарной коры кошки.// Сенс. сист., 2001, т. 15, №1, с.29-35
Глезер В.Д., Праздникова Н.В., Леушина Л.И., Павловская М.Б. Опознание зрительных образов // Гл. IX в Руководстве по физиологии, т. Физиология зрения. М. Изд-во Российской АН 1992, с. 466-527.
Глезер В.Д., Невская А.А., Гаузельман В.Е., Чернова Н.Д. Выделение фигуры из фона: межполушарные различия и вероятная локализация механизмов. // Сенс. сист., 2002, т. 16, №2.
Леушина Л.И. Зрительное пространственное восприятие Л., 1978, 175 с.
Леушина Л.И., Невская А.А. Анализ экспериментального распределения ответов, - выделение стимулов, обусловленных стимулом. // кн. Леушина Л.И. Зрительное пространственное восприятие Л., 1978 , с 143-151.
Леушина Л.И., Невская А.А., Павловская М.Б. Асимметрия полушарий головного мозга с точки зрения опознания зрительных образов. // В кн. Сенсорные системы: Зрение. Л., 1982, с. 76 – 92.
Невская А.А. Межполушарные различия при зрительном восприятии: спорные вопросы и переспективы исследований. // В. кн. : Сенсорные процессы и асимметрия полушарий. Л., 1985, с. 3-21.
Невская А.А., Леушина Л.И. Асимметрия полушарий и опознание зрительных образов. Л., 1990. с.152
Невская А.А., Леушина Л.И. Особенности использования гармонического пространственно-частотного спектра изображений при их описании в правом и левом полушариях. // Сенс. сист., 1995, т. 9, № 1, с. 108 – 116.
Праздникова Н.В., Глезер В.Д., Макаров Ф.Н. О механизмах инвариантного описания образа и полного описания изображения в левом и правом полушариях кошки. // Сенс. сист., 1997, т. 11, № 3, с 352 – 359.
Чернова Н.Д., Невская А.А., Глезер В.Д., Гаузельман В.Е. Роль взаимодействия полушарий в опознании зрительных образов. Сенс. сист., 2000, т. 14, № 2, с 167 – 172.
Bryden M.P., Allard F. Visual hemifield differences depend on typeface. // Brain a Lannguage, 1976, vol. 3, p. 191 – 200.
Franco L., Sperry R.W. Hemisphere lateralization for cognitive processing of geometry. // Neuropsyhologia, 1977, vol. 15, p. 107-114.
Fu K. (Фу К.) Структурные методы в распознавании образов. М., 1977, 319 с.
Gazzanda M.S., Bogen J.E., Sperry R.W. Observations of visual perception after disconnection of the cerebral hemispheres in man. // Drain, 1965, vol. 88, p. 221-236.
Hebb D.O. The organization of behavior. New York, Wiley, 1949, 335 p.
Hecaen H. Aphasic, apraxic and agnostic syndromes in right and left hemisphere lesions. // Handbook of clinical neurobody., vol. 4, Amsterdam, 1969, p. 291-311.
Hunt E. (Хант Э.) Искусственный интеллект. М., 1978, 558 с.
Levy J., Trevarthen C., Sperry R.W. Perception on bilateral chimeric figures following hemispheric deconnection. // Brain, 1972, vol. 95, p. 61-78.
Marcolec C.J. Abstract visual form representations in the left cerebral hemisphere. // J. Exper. Psychol.: Human Percept. A. Perform., 1955, vol. 21, p. 375-386/
Moscovitch M. Information processing and the cerebral hemispheres. // In: The handbook of behavioral neurobiology., vol. 2, Ed. By M.S. Gazzaniga, New York, 1979, p. 379-446/
Sperry R.W., Zeidel E., Zeidel D. Self recognition and social awareness in the deconnected minor hemisphere. // Neuropsychologia, 1979, vol. 17