ФОРМИРОВАНИЕ ФУНКЦИОНАЛЬНОЙ МЕЖПОЛУШАРНОЙ АСИММЕТРИИ МОЗГА В ДИНАМИКЕ ОБУЧЕНИЯ
Г.А. Кураев, И.В. Соболева, Л.Г. Сороколетова
Ростовский Государственный Университет
Проблема симметрии и асимметрии в природе принадлежит к числу фундаментальных естественно-научных и философских проблем [1, 82, 74, 76, 92].
В эволюции любая биологическая система стремится к устойчивому состоянию, а следовательно, к симметрии. Однако внутри таких систем возникают процессы, нарушающие симметрию, что способствует дальнейшему развитию, и в то же время, к перестройке ранее сформировавшейся системы. Наиболее наглядно это выявляется при анализе функциональной организации процессов, происходящих в центральной нервной системе, построенной по билатеральному типу.
Взаимоотношения симметричных структур больших полушарий головного мозга привлекают внимание нейрофизиологов в связи с расширившимися исследованиями роли межполушарных отношений в таких интегративных процессах, как замыкание временной связи, хранение следов обучения, пространственный анализ, организация эмоциональной окраски поведения, компенсаторные процессы, распознавание образов [2, 16, 17, 89, 90, 71, 3, 25 и др.].
В эволюционных перестройках головного мозга особое значение имеет прогрессирующее функциональное и морфологическое дифференцирование корковых полей, усиление комиссуральных межполушарных систем, на основе чего происходит функциональная доминирующая специализация симметричных структур мозга, венцом которой является специализация полушарий у человека. Накопленные в физиологии данные свидетельствуют о том, что функциональная асимметрия полушарий головного мозга имеет место и у животных [18, 19, 72, 58].
На исследования симметрии и асимметрии больших полушарий головного мозга, особенно за последние годы, обращено пристальное внимание специалистов разного профиля, работающих в области общей биологии, медицины, нейроморфологии, нейрофизиологии, иммунологии, спорта. В теоретическом плане такие исследования необходимы для познания принципов организации и механизмов совместной деятельности симметричных структур мозга. В практическом отношении раскрытие механизмов межполушарных отношений необходимо для решения ряда вопросов, связанных с клиникой и патогенезом некоторых форм заболеваний мозга, с проблемой управления функциональным состоянием организма и разработкой новых рациональных методов оптимизации обучения [6, 20, 100, 9, 71, 45, 92, 25, 3, 62].
Большой фактический материал, накопленный в исследованиях симметричных структур головного мозга, хотя и дает представление о некоторых механизмах их взаимодействия, тем не менее до настоящего времени оставляет нерешенным фундаментальный вопрос о функциональном значении симметричного строения высших отделов головного мозга в организации различных форм поведения.
Динамика межполушарных отношений в процессе образования условного рефлекса
С тех пор как Broca [106] установил факт о левополушарной локализации нервных структур, организующих речевую функцию, на длительное время утвердилось мнение о тотальном доминировании левого полушария и в других функциях у человека. Причем, наличие полушарной специализации считалось особенностью эволюционного развития, присущей только человеческому мозгу. В дальнейшем рядом исследователей была обоснована точка зрения о неправомерности такого заключения.
В.Л. Бианки [19] отмечен ряд факторов, явившихся причиной отрицания распределения функций между полушариями у животных: равная вероятность встречаемости у них правшества и левшества; отсутствие различий в анализе сигналов левым и правым полушариями в опытах с расщепленным мозгом. Автор вполне справедливо замечает, что причиной таких результатов были: а) ослабление исходной асимметрии при расщеплении мозга; б) опыты с повреждением симметричных зон коры не могли быть абсолютно идентичными по степени повреждения, развитию отечности после операции и т.д., поэтому, такие исследования не могут служить основой для выяснения вопроса о наличии асимметрии у животных; в) отсутствие в большинстве исследований учета пола животного. К сказанному можно добавить, что большое значение для выявления асимметрии имеет адекватность выбранного стимула. На наш взгляд, прежде всего, этот факт следует учитывать при попытке выявить межполушарную асимметрию в разных зонах коры.
Существует множество методических подходов исследования межполушарной асимметрии мозга [2, 65, 16, 67, 25] из которых можно сделать вывод, что наиболее успешно функциональная организация симметричных структур мозга раскрывается при изучении ее в динамике становления какой либо из форм поведения.
В наших экспериментах по электрофизиологическим и поведенческим коррелятам прослежены отношения симметричных систем коры мозга в исходном состоянии и их качественные перестройки в ходе выработки условных рефлексов.
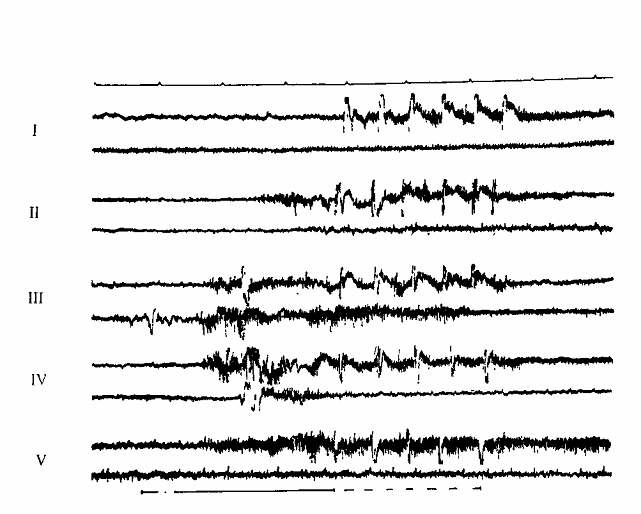
Уже по показателями поведенческих условно-рефлекторных двигательных (оборонительных и пищевых) реакций на сигнал стало ясно, что межполушарное взаимодействие в процессе формирования рефлекса характеризуется сменой функциональных состояний симметрии и асимметрии. Внешне это проявилось в определенной динамике двигательных реакций на сигнал, что позволило нам выделить 5 этапов взаимодействия полушарий головного мозга при выработке оборонительного условного рефлекса (рис.1).
На первом этапе сигнал еще не вызывал условной двигательной реакции, ответ был только на подкрепление, что хорошо иллюстрируется активностью миограммы рисунка 1. На втором этапе на стороне подкрепления появлялись локальные двигательные реакции на сигнал. Видимо, на этом этапе в организации условного рефлекса доминирующую роль играло то полушарие мозга, в которое преимущественно адресовалось подкрепляющее раздражение. На этом этапе генерализация условного рефлекса еще не охватывала симметричные пункты второго полушария. На 3-м и 4-м этапах двигательные реакции на сигнал были двусторонними, но на 3-м этапе они были синергичными, а на 4-м – реципрокными. Различия в характере эффекторных реакций на 3-м и 4-м этапах, видимо, обусловлены тем, что на 3-м этапе выработки рефлекса возбуждение одновременно охватывает моторные центры полушарий, обеспечивающие флексорную реакцию конечности. Это приводит к торможению, на основе индукционных отношений, центров, обеспечивающих экстензию конечности. На 4-м этапе, вследствие усиливающегося тормозного влияния со стороны доминирующего полушария, флексорный центр в субдоминантном полушарии тормозится, а это приводит к временному возбуждению экстензорного центра, что и проявляется реципрокностью двигательных реакций на сигнал.
Рис. 1. Миограммы мышц конечностей на I-V этапах выработки рефлекса. Верхние записи в каждой паре - регистрация от мышц конечности, на которую наносилось подкрепляющее раздражение, нижние - от мышц симметричных конечностей. Сплошная линия-время действия сигнала, пунктир - время действия подкрепления Отметка времени - I с.
На пятом этапе выработки рефлекса сигнал вызывал локальные условные реакции только на стороне подкрепления. Следовательно, на этом этапе генерализация сменялась концентрацией нервных процессов в полушарии, организующем условно-рефлекторный ответ. Такая динамика наблюдалась при выработке оборонительного условного рефлекса с применением в качестве подкрепления электрокожного раздражения лапы или уха животного.
Наличие межполушарной генерализации условно-рефлекторной деятельности проявилось и в опытах с выработкой пищевого условного рефлекса в условиях свободного выбора лапы для сигнальной инструментальной реакции, которую животные выполняли вначале только доминирующей лапой, затем доминирующей и субдоминантной (с одинаковой вероятностью). Лишь после этого наступало упрочение рефлекса с четкой односторонней реакцией на сигнал (Рис.2).
Такая динамика в использовании лапы для инструментальной реакции свидетельствует о динамике активности полушарий от асимметричного их состояния к симметричному и вновь к асимметричному [58, 24].
В наших экспериментах с выработкой пищевого инструментального условного рефлекса в условиях, когда животное имело возможность свободного выбора конечности для манипуляции (нажим на рычаг) в ответ на сигнал, по мере упрочения рефлекса «выбиралась» одна и та же лапа, независимо от латерализации рычага относительно животного, движущегося к кормушке. При этом, в случаях, когда рычаг был расположен на одной стороне с доминирующей лапой, скорость упрочения рефлекса была большей, чем при разноименной латерализации.
Выбор животным лапы для инструментального воздействия можно расценить как свидетельство наличия у кошек исходной функциональной асимметрии полушарий.
В другой постановке опыта, с выработкой оборонительного условного рефлекса оказалось, что время условно-рефлекторных процессов, необходимое для упрочения рефлекса (о чем можно судить по количеству сочетаний, предшествовавших исчезновению условной реакции на нераздражаемой стороне), зависело от совпадения или несовпадения стороны подкрепления с доминирующей лапой. Так, у кошек-правшей при нанесении подкрепления на правую лапу для исчезновения условной реакции, неподкрепляемой конечности в среднем требовалось около 260 сочетаний. В
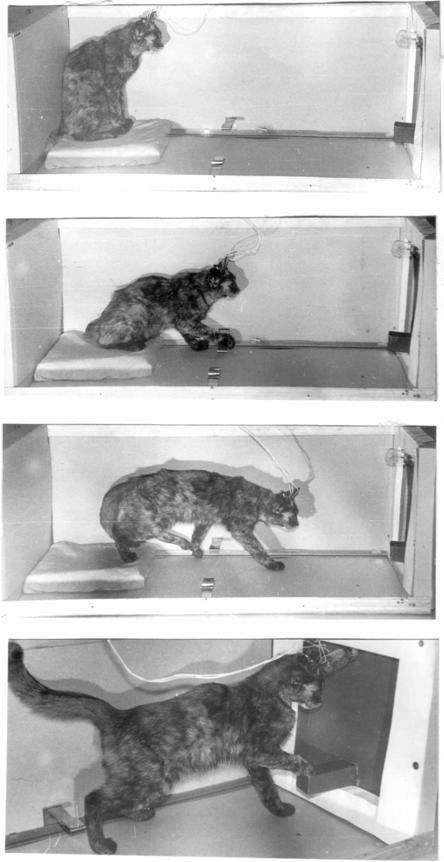
Рис.2. Динамика выработка условного инструментального рефлекса у кошки. Пояснения в тексте.
тех же случаях, когда у правшей подкрепление наносилось на левую лапу, среднее число сочетаний, необходимое для аналогичного результата, было более 400 сочетаний. Это, видимо, обусловлено тем, что доминантность полушария, необходимая для организации условной реакции, формировалась в конкурентной борьбе с исходной, врожденной доминантностью коры противоположного полушария.
Возможно, что за счет преимущественной афферентации в полушарие, доминирующее по организации двигательной активности животного, ускоряется концентрация нервных процессов после их генерализации. По всей вероятности, это происходит благодаря усилению тормозных влияний со стороны доминантного полушария на субдоминантное.
Таким образом, на основе анализа динамики образования условного оборонительного и пищевого рефлексов, можно выделить в межполушарных отношениях пять этапов межполушарного взаимодействия. Время генерализации и концентрации условного рефлекса зависит от преимущественной адресации подкрепляющего воздействия в доминантное или субдоминантное полушарие. В первом случае скорость выработки условного рефлекса выше, исходная межполушарная асимметрия усиливается. Во втором – скорость выработки условного рефлекса ниже, исходная межполушарная асимметрия для данной двигательной реакции инвертируется.
Особенности функциональной организации симметричныхпунктов зрительной и соматосенсорной коры
Выявленная в поведенческих опытах динамика межполушарных отношений при формировании условно-рефлекторных двигательных реакций, зависимость их от совпадения стороны подкрепляющего раздражения с доминирующей конечностью, ставят вопрос о существовании различий полушарной активности априорно, т.е. в фоновом и вызванном режимах.
Наши исследования суммарной и импульсной активности показали, что в фоновой электрокортикограмме (ЭкоГ), регистрируемой в симметричных пунктах зрительной и соматосенсорной коры полушарий более высокочастотные ритмы преобладали в полушарии, контралатеральном относительно ведущей конечности. Эта зависимость была достоверной для соматосенсорной коры и в форме тенденции наблюдалась в зрительной коре.
Асимметрия волн медленных ритмов ЭкоГ [32] в сенсомоторной коре доминирующего по моторике полушария, как правило, была менее выражена и чаще имела отрицательный знак, что можно расценить как преобладание в этом полушарии возбудительных процессов.
У большинства животных в субдоминантном полушарии было больше нейронов с пачечной и групповой формами активности, а в доминантном с тонической (рис.3).
Средняя частота импульсных разрядов была больше у нейронов, зарегистрированных в соматосенсорной коре доминирующего полушария. Кроме того, нейронная активность доминирующего полушария имела большую вариабельность и меньшую скоррелированность импульсных потоков соседних нейронов.
Выявленные отличия в наших исследованиях по фоновой активности доминирующего и субдоминантного полушарий подтвердились исследованиями с применением световой и электрокожной стимуляций.
Так, в ЭкоГ зрительной коры световая стимуляция усиливала выраженность быстрых ритмов глубоких слоев, преимущественно в полушарии, доминирующем по лапе.
Межполушарные отличия однотипных зон коры отмечены и при анализе фокальной активности. Световое раздражение вызывало в 17-м поле доминирующего полушария достоверно более коротколатентные (сравнительно с субдоминантным) фокальные ответы. В 18-м поле, наоборот, коротколатентные ответы были в субдоминантном полушарии. Неадекватные стимулы, хотя и вызывали изменения фокальной активности коры, но они были однотипны в обоих полушариях.
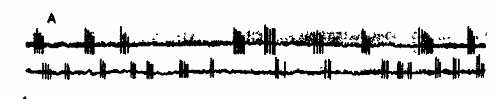
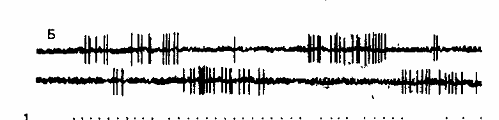

Рис. 3 . Тип фоновой активности и реакций нейронов»
А — пачечные; Б — групповые; В — тонические нейроны; I — отметка времени. Стрелкой отмечен момент нанесения раздражения.
В 17-м поле зрительной и соматосенсорной коры доминантного полушария реагирующих нейронов было больше (за счет возбуждающихся), латентные периоды их реакций были короче, вариабельность средней частоты потоков импульсов ниже, чем в субдоминантном. В 18-м поле зрительной коры превосходящее число реагирующих нейронов отмечено в субдоминантном полушарии. В нем же была несколько большая средняя частота разрядов нейронов, короче ЛП реакций, однако, вариабельность средней частоты была выше в доминантном полушарии.
В 17-м и 18-м полях зрительной коры количество полисенсорных нейронов превалировало в доминантном полушарии. Точно также, в соматосенсорной коре у кошек-правшей в левом, а у кошек-левшей в правом полушарии бисенсорных нейронов больше, чем в симметричных пунктах противоположного полушария.
При неадекватной стимуляции (свет для соматосенсорной, ЭКР для зрительной) нейрональная активность симметричных пунктов коры, как правило, мало отличалась. В этом, видимо, проявляется эволюционное развитие анализаторных областей коры, заключающееся в разделении ее на зоны с преобладанием элементов, настроенных реагировать на специфические раздражители (моносенсорных нейронов), и зоны с преобладанием элементов ассоциативного типа (полисенсорных нейронов). Одновременно с этим происходит дифференциация функций между полушариями.
Нашими опытами установлено, что функциональное доминирование возникает лишь в случае, если применяется раздражение, адекватное по своей сенсорности для коркового конца данного анализатора. Так, применение светового раздражения сопровождалось межполушарной асимметрией только зрительных зон коры, а применение электрокожного – соматосенсорных.
В динамике образования оборонительного условного рефлекса, что прослеживалось по изменениям суммарной, фокальной и импульсной активности межполушарные различия этих показателей были существенно иными, чем при действии индифферентных стимулов [55, 60]. Это проявилось в том, что в динамике образования условного рефлекса корковые процессы имеют специфические особенности на разных этапах его формирования [48].
Т.С. Наумовой [73] выделено пять стадий изменений электрической активности коры при формировании условного рефлекса. Первые две из них характеризовались диффузными изменениями ЭЭГ, проявляющимися и при воздействии индифферентными стимулами, либо в форме депрессии электрической активности, либо в форме ее экзальтации. Стадия генерализации, согласно Т.С. Наумовой [73], продолжалась у разных животных в течение 100-110 сочетаний, что в наших исследованиях соответствовало первым этапам выработки рефлекса. Поверхностные электроды, которые использовались указанным автором, как справедливо отмечал М.Я. Рабинович [80], слабо отражают быстрые процессы глубоких структур, поэтому при упроченном рефлексе в ЭкоГ-активности не удавалось отметить каких-либо изменений. Такое представление согласуется с результатами работы И.Н. Книпст [48], установившей при выработке условного рефлекса в зонах проекции сигнального и подкрепляющего раздражений определенную динамику усвоения ритма условной стимуляции от верхних слоев коры к нижним.
Как уже отмечалось выше, на основании анализа опытов с выработкой оборонительного и пищевого рефлексов нами было выделено пять этапов становления условного рефлекса. Эти этапы определялись, как уже говорилось, не по характеру электрической активности, а по динамике условной двигательной реакции симметрично расположенных мышечных групп. Однако, упрочение условного рефлекса имело на этих этапах и специфические особенности, отражаемые в активности корковых процессов.
На первом этапе, когда двигательная реакция возникала только на безусловный стимул, в зрительной коре верхних и нижних слоев контралатерального относительно подкрепления полушария в последействии сочетаний усиливалась медленная ритмика. В предсигнальный, сигнальный периоды и период подкрепления полушарных отличий в суммарной активности не было (рис.4).
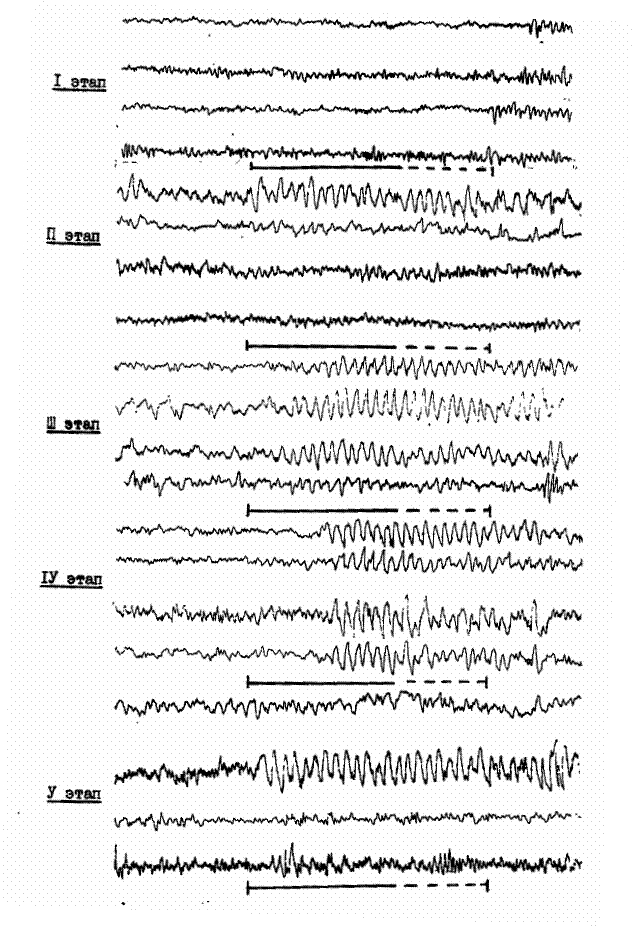
Рис. 4. Динамика электрокортикограммы симметричных пунктов зрительной коры мозга кошки на разных этапах выработки условного рефлекса. Внизу: отметка условного (сплошная линия) и безусловного раздражителей.
Несколько иная ситуация наблюдалась на этом этапе в соматосенсорной коре. Оказалось, что здесь в предсигнальный и сигнальный периоды межполушарных отличий, как и в зрительной коре, не было. Однако в контралатеральном полушарии выраженность тэта-ритма и возбудимость коры усиливались в период подкрепления в глубоких слоях, а в период последействия – в верхних.
На втором этапе, когда стали проявляться первые односторонние условные реакции, в ЭкоГ зрительной коры, регистрируемой на всех периодах условного рефлекса (предсигнальный, сочетания и последействия) выраженность медленной ритмики увеличивалась, причем больше в контралатеральном полушарии. Увеличивалась и возбудимость нижних слоев коры. Для этого этапа, особенно в период подкрепления, было характерно увеличение межполушарной асинхронности ритмов медленной активности.
В соматосенсорной коре в предсигнальный и сигнальный периоды медленные ритмы преобладали в верхних слоях обоих полушарий, а в период последействия во всех слоях коры, при этом после предъявления сочетаний послойные различия в ЭкоГ исчезали. Следует отметить, что в большинстве случаев медленная ритмика преобладала в контралатеральном полушарии. Асинхронность межполушарной активности между глубокими слоями коры отмечалась в основном в период действия сигнала и подкрепления.
Наметившиеся на втором этапе выработки рефлекса различия полушарий в функциональной организации зрительной и соматосенсорной коры не получали дальнейшего развития на третьем этапе, когда возникала билатеральная реакция на сигнал. На этом этапе близкие функциональные состояния в обоих полушариях выявились в форме повышения сходства активности и возбудимости всех слоев коры, в том числе в форме сниженной межполушарной асинхронности медленных ритмов.
На четвертом этапе, когда появились реципрокные двигательные реакции на сигнал, сходство состояний активности симметричных пунктов зрительной коры сохранялось, но это происходило на фоне уменьшения в верхних слоях выраженности медленных ритмов и ее усиления в нижних, параллельно с этим в сигнальный и предсигнальный периоды происходило усиление возбудимости коры. В те же периоды отмечалась и межполушарная функциональная асимметрия, что отразилось в асинхронности возникновения в полушариях медленных ритмов. В соматосенсорной коре на данном этапе выработки рефлекса продолжала сохраняться одинаковая выраженность медленных ритмов во всех слоях коры обоих полушарий в предсигнальный период и в период действия сигнала. При подкреплении и после сочетания высокие значения тэта-индекса сохранялись только в глубоких слоях коры контралатерального полушария. Большая возбудимость коры была, как правило, в контралатеральном полушарии. На этом фоне лучшая выраженность межполушарной асинхронности развития медленных ритмов проявлялась в предсигнальный и сигнальный периоды. Следовательно, на этом этапе как в зрительной, так и в соматосенсорной коре начинает вновь формироваться межполушарная асимметрия корковых процессов.
Наиболее четкие межполушарные отличия в характере суммарной активности наблюдались на пятом этапе выработки условного оборонительного рефлекса (при локальной односторонней реакции на сигнал) между глубокими слоями коры. В период действия сигнала в этих слоях сохранялась высокая возбудимость и выраженность тэта-ритма, а в остальных слоях и в ипсилатеральном полушарии, активность становилась близкой к фоновой до выработки рефлекса. Данная динамика выраженности тэта-ритма в симметричных пунктах полушарий соответствует фазам генерализации и концентрации условного рефлекса. При упроченном рефлексе сохранение сигнальных перестроек обычно отмечается только в тех пунктах, которые расположены в зонах проекции анализаторов, участвующих в осуществлении условного рефлекса [63, 56].
Таким образом, анализ суммарной активности показал, что усиление тэта-ритма и усвоение ритма в динамике выработки рефлекса вначале наиболее выраженно происходит в верхних слоях коры контралатерального относительно стороны подкрепления полушария, а затем в тех же слоях ипсилатерального, глубоких слоях контралатерального, и, наконец, ипсилатерального полушария. При упроченном рефлексе наибольшие изменения ЭкоГ-активности сохраняются в глубоких слоях коры контралатерального полушария. Такая динамика имела место в разные периоды: в предсигнальный, сигнальный, подкрепления и последействия. Однако, время появления таких изменений от начала выработки рефлекса для всех периодов не было одинаковым.
Раньше всего они возникали в последействии, затем в периоды подкрепления и действия сигнала и, наконец, в предсигнальный. Следовательно, начальные изменения активности происходят на следах предшествующих сочетаний [73] и это характерно для обоих полушарий. В отмеченной последовательности изменялась и возбудимость коры, о чем можно судить по динамике асимметрии волн тэта-ритма в ходе выработки рефлекса. Сходная динамика выявлена при анализе асинхронности корковой активности симметричных областей мозга.
Исследования, проведенные с применением регистрации фокального потенциала, принципиально подтвердили динамику развития межполушарного взаимодействия, выявленную по ЭкоГ-показателям, но при этом определились некоторые особенности динамики возбудительных и тормозных процессов. В качестве критерия выработки рефлекса нами было использовано появление в фокальном ответе позднего позитивного колебания (Рис.5).
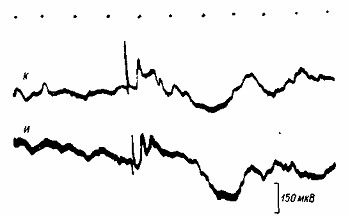
Рис.5. Фокальные вызванные потенциалы симметричных пунктов поля 18 зрительной коры. И — ипси-, К — контралатеральное полушарие относительно стороны подкрепляющего раздражения. Отметка времени — 50 мс. Стрелки — момент предъявления вспышек. Отметка времени — 50 мс.
На первом этапе выработки рефлекса в фокальных ответах поздний позитивный потенциал на вспышки света, используемые как сигнал, не встречался. Первичные негативные колебания в разных полушариях имели однотипные величины латентных периодов и длительностей. По мере упрочения рефлекса появлялись различия в характере ответов, регистрируемых в разных полушариях: в ипсилатеральном – длительность первичных негативных колебаний, латентные периоды и их вариабельность становились больше (рис.6).
Поздний позитивный потенциал, появление которого свидетельствует об усилении тормозных процессов, на первых этапах выработки рефлекса был мало отличим в обоих полушариях, но количественно в ответах на повторяющиеся вспышки света он преобладал в полушарии, контралатеральном относительно подкрепления. Следовательно, тормозные явления в последнем были более интенсивны.
Начиная с третьего этапа, межполушарные различия проявлялись по характеру поздних позитивных колебаний. В ипсилатеральном полушарии ЛП и длительность позитивных волн были больше, чем в контралатеральном. Следовательно, лабильность процессов в последнем была выше.
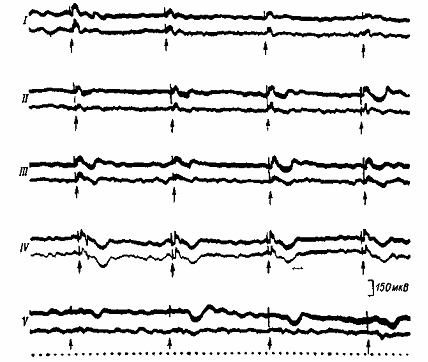
Рис. 6. Фокальные ответы симметричных пунктов поля 18 зрительной коры. Римские цифры — этапы выработки условного рефлекса. Стрелки — момент предъявления вспышки света. Верхние осциллограммы в каждой паре — запись из коры контралате-рального полушария, нижние — ипсилатерального. Отметка времени — 50 мс.
Повторение сочетаний на четвертом и пятом этапах приводило к резкому урежению появления поздних позитивных колебаний в фокальных ответах, регистрируемых из ипсилатерального полушария. Скорость появления этих колебаний в контралатеральном полушарии и урежение их в ипсилатеральном были выше, если латеральность подкрепления совпадала с доминирующей по лапе стороной, и наоборот.
В динамике выработки рефлекса изменялся рисунок позднего позитивного колебания: передний фронт его удлинялся, задний – укорачивался. Эти изменения в основном отмечались в полушарии, контралатеральном относительно подкрепления.
Таким образом, в динамике выработки условного рефлекса поздний позитивный потенциал вначале появляется в контралатеральном полушарии, затем в ипсилатеральном, что в итоге приводит к однотипности характера активности обоих полушарий, а на пятом этапе он вызывается только в контралатеральном. На пятом этапе число сигнальных вспышек, сопровождающихся поздними позитивными колебаниями, уменьшается. Так как эти колебания свидетельствуют об усилении тормозных процессов, то, следовательно, по мере выработки рефлекса торможение вначале растет в контралатеральном, а затем ипсилатеральном полушариях; при упроченном же рефлексе оно более выражено в контралатеральном.
Анализ нейронной активности, регистрируемой в симметричных пунктах зрительной и соматосенсорной коры, показал, что условно-рефлекторное обучение сопровождается определенным изменением последовательности разрядов отдельных нейронов. Эти изменения в разных полушариях были выражены неоднозначно. Прежде чем обсудить межполушарные отличия нейронной активности в динамике выработки оборонительного рефлекса, остановимся на некоторых общих моментах.
Хорошо известно, что в динамике выработки условного рефлекса уменьшается или полностью исчезает условная активность нейронов сигнальной зоны коры [104]. Однако, исчезновение реакции нейронов в форме изменения частоты разрядов (признак, на который опирается большинство авторов) не может служить доказательством прекращения участия нейронов в условно-рефлекторном процессе. Примененный нами метод угашения рефлекса и наблюдаемые при этом изменения реактивности нейронов доказывает, что условно-рефлекторная реакция при упроченном рефлексе обеспечивается иным, чем в процессе выработки, способом.
В работах Doty [109], Э.Г. Заркешева [41, 42] указывается, что в любой структуре мозга имеются обучающиеся клетки, реализующие замыкание данного вида временной связи. В нашем исследовании нейроны, усиливающие реакцию на сигнал в начале угашения рефлекса были отнесены к «обучающимся», а неизменявшие – к «необучающимся». У «обучающихся» нейронов в динамике выработки условного рефлекса независимо от зоны регистрации (первичная и вторичная зрительная, соматосенсорная кора), вариабельность частоты разрядов уменьшалась в фоне при предъявлении сочетаний и в последействии. Это, по-видимому, связано с возрастанием упорядоченности работы систем нейронов, участвующих в условно-рефлекторном процессе. Параллельно этому увеличивалась корреляция потоков импульсных разрядов нейронов, зарегистрированных одновременно по одному каналу. «Обучающиеся» нейроны реагировали на сигнальные стимулы с более длинными латентными периодами. В структуре их импульсного потока (в динамике выработки условного рефлекса), как правило, формировалась тормозная пауза, которая по времени строго соответствовала позитивному колебанию фокального ответа, регистрируемого тем же электродом, что и импульсный разряд.
Для «необучающихся» нейронов указанные закономерности как при выработке рефлекса, так и при его угашении не были характерны.
Итак, на основании нейрофизиологических исследований можно считать, что в динамике выработки оборонительного условного рефлекса в зонах сигнала и подкрепления характер активности зависит от участия или неучастия их в условно-рефлекторном процессе [27, 102, 103]. Так, если для «необучающихся» нейронов было характерно изменение степени активности – вначале усиление с последующим снижением ее до исходного уровня, то у большинства «обучающихся» на первых этапах выработки рефлекса частота разрядов в ответах на сигнал также увеличивалась, а затем их разряды приобретали особый порядок следования импульсов.
Это, по-видимому, обусловлено тем, что центральные нейроны используют разные коды и могут при обучении приобретать новые способы кодирования, обучаться им. Последнее доказывается тем, что нейроны при образовании временной связи изменяют свою активность соответственно реакции, вызванной подкрепляющим стимулом. Здесь следует оговориться относительно нейронов коры, детектирующих специфические стимулы. Эти нейроны отличаются низкой способностью к пластическим изменениям своей активности. Качественные перестройки в функционировании нейронов коры при обучении, возможность одним нейроном использовать разные коды, его способность обучаться новым кодам имеет большое значение для объяснения высокой пластичности нервной системы.
Способы кодирования не одинаково распространены на различных уровнях центральной нервной системы. Частотное кодирование интенсивности стимула имеет место преимущественно в ее периферических отделах и прежде всего в рецептирующих структурах [77]. На этом уровне обнаружено кодирование вероятностью ответа, латентностью [99]. Признавая такие способы. Г.Г. Вайткявичус и др., [28] сообщают о возможности кодирования в периферических рецепторах номером канала, что обеспечивает селективность реакций нейрона на определенную интенсивность раздражения.
По данным Б.Я. Пятигорского [79], уже на рецепторном уровне при наличии контроля его работы со стороны других структур или конвергенции импульсов на одном элементе, кодирование частотой меняется на более сложный интервальный код.
По-видимому, в центральной нервной системе наблюдается кодирование преимущественно структурой потока импульсов с различным чередованием межимпульсных интервалов. Это доказывается рядом работ. Так, Segundo et al., [122] выявлено, что сила сокращения мышечного препарата зависит от характера распределения коротких и длинных интервалов импульсации, идущей из центра. П.Г. Костюком [54] показана способность мотонейронов спинного мозга к выделению физических компонентов разрядов в афферентных потоках; В.Д. Глезер и др., [33] установили наличие кодирования группами разрядов в центральных отделах зрительного анализатора позвоночных.
При такой сложной форме поведения, как условно-рефлекторное, выявлено, что нейроны зоны корковой проекции условного раздражения используют частотный код, но при этом повышается значимость паттерна межимпульсных интервалов [29, 85].
Исследования Gerstein et al. [110], связанные с изучением временной структуры возбуждения нейронов слуховой коры как функции параметра стимула, показали, что, хотя каждый нейрон на многие предъявления стимула отвечал вариабельно, в его ответах имелся статистически значимо повторяемый паттерн импульсной последовательности. Возможность кодирования паттерном импульсов доказывается тем, что полимодальные нейроны могут отвечать разными паттернами на разные стимулы. К этому следует добавить, что характерные паттерны возбуждения в обширной зоне коры на один и тот же условный стимул были одинаковыми, в то же время в одних и тех же пунктах коры на два отличающихся стимула возникали существенно различные паттерны возбуждения, причем во всех случаях интенсивность реакции изменялась незначительно.
Все приведенные факты свидетельствуют в пользу наличия в центральной нервной системе кодирования информации узором импульсного потока, величинами межимпульсного интервала. В то же время по паттерну возбуждение одиночного нейрона невозможно предсказать, какова будет двигательная или иная поведенческая реакция животного. Видимо, обработка информации для организации поведения опосредуется одновременной активностью ансамблей нейронов, имеющих определенные границы и взаимоотношения, которые варьируют в зависимости от характера выполняемой задачи и других факторов.
По представлению А.М. Иваницкого [43], в нервной системе специфичность сигнала сохраняется с помощью определенной архитектуры нервной связи, особенности нервного пути. Поэтому один и тот же импульс имеет разное значение в зависимости от того, по какому пути он пришел к нервному центру или клетке. При телеграфной связи такой способ потребовал бы всего 32 линии – по одной для каждой буквы. Это, с точки зрения автора, освобождает нервный импульс от нагрузки нести «смысловую информацию». Следовательно, данное представление полностью исключает участие самой нервной клетки в переработке информации и отводит ей только роль в переориентации пути импульсного потока. Таким образом, специфика нервной информации определяется не последовательностью импульсов, не количеством разрядов в единицу времени, не величиной импульсного интервала, а местом нейрона в сложной системе нейрональных связей. Однако, следует учитывать, что нервная клетка не столь уж индифферентна к значению сигнала, о чем свидетельствуют данные Р.С. Мнухиной и др. [70], показывающие, что при замыкании временной связи происходят изменения длительности потенциалов действия нейрона.
Известен ряд работ, свидетельствующих о большой пластичности нейрональных реакций при обучении. В то же время, как уже отмечалось, исследования нейрональных механизмов обучения по электрическим проявлениям показали, что по мере становления и упрочения рефлекса реакции фоновой ритмики, а также вызванные потенциалы и импульсная активность, сопровождающие условно-рефлекторную реакцию, ярко выраженные вначале, на более поздних стадиях ослабевают или не выявляются вовсе [29]. Это может быть связано как с реализацией выработанного условного рефлекса лишь небольшим числом клеточных элементов, так и обеспечением условно-рефлекторных реакций скорее перестройками узора импульсного потока, чем существенными изменениями его интенсивности.
Исследования в опытах с оборонительным условным рефлексом, динамики его становления, позволили нам предположить, что значительная часть нейронов для организации условной реакции вначале использует частотный код, который по мере упрочения условного рефлекса не выявляется. Однако, анализ узора потока межимпульсных интервалов позволил заметить стабильное увеличение частоты встречаемости некоторых из них – определенных для данного нейрона.
В.Л. Силаковым [85] выдвинуто положение о существовании в зрительной коре двух типов дистрибутивного кодирования: пачечного и группового. Наши данные [58] показывают, что нейроны с такими формами активности встречаются в фоне в относительно малом числе случаев, но в динамике выработки рефлекса встречаемость таких нейронов возрастала. Причем больше таких нейронов было в 18 поле, сравнительно с 17 полем, зрительной коры. Следовательно, пачечная и групповая форма активности нейронов являются одними из форм кодирования сигнальной информации при условно-рефлекторной деятельности. Тем более, что, по мнению того же В.Л. Силакова [85], переход на пачечный режим обеспечивает высокую надежность передачи информации, поскольку эффективность пачки импульсов выше, а вероятность случайной блокады ее ниже, чем одиночных разрядов.
При выработке рефлекса повышается сходство активности по характеру межимпульсных интервалов в фоне с интервалами в момент условной реакции. Это свидетельствует, что в нейрональной системе происходят существенные перестройки на оптимальное решение задачи до предъявления пускового сигнала. В.Л. Силаков [85], Н.П. Бехтерева [15] предполагают, что это связано с генерализацией паттернов активности, связанных с обуславливанием и отражающих этап фиксации следов условного сигнала. Следовательно, определенный межимпульсный интервал, доминирующий в период условной стимуляции, при выработке рефлекса имеет место в фоновой активности, предшествующей условному стимулу. В наших исследованиях оказалось, что в случаях, когда животное проявляло условную реакцию в межсигнальный период, характер распределения межимпульсных интервалов активности нейронов был сходен с выявленным при условной стимуляции. Однако, чаще количественная выраженность специфического узора межимпульсных интервалов в фоне была незначительной и увеличивалась при предъявлении условного стимула. Следовательно, для вызова условного рефлекса необходимо оптимальное число определенных межимпульсных интервалов. Это, естественно, связывает условную реакцию с некоторыми изменениями частоты импульсации. Кодирование условного сигнала, по-видимому, осуществляется двояко: узором межимпульсных интервалов и частотой разрядов. Вес каждого из этих способов на разных этапах образования условного рефлекса и его реализации различен. По нашим данным перестройки импульсной активности нейрона в динамике обучения сопровождаются превалированием вначале изменений частоты импульсации, а затем проявлением особого паттерна межимпульсных интервалов. По данным Н.Н. Василевского [29], кодирование интенсивностью потока используется для срочной, надежной, но грубой передачи информации, а упорядоченными потоками – для точной ее передачи.
Сравнение динамики межимпульсных интервалов потоков активности нейронов симметричных пунктов коры показало, что при выработке оборонительного условного рефлекса перестройки характера импульсной активности раньше происходят в коре контралатерального полушария, затем появляются в коре ипсилатерального. На этапе упроченного рефлекса, когда имела место изолированная односторонняя двигательная реакция, в ответ на сигнал специфический узор возникал у нейронов только этого полушария (рис.7).
Видимо, в обоих полушариях способы кодирования сигнальной информации одинаковы, но на стадии упроченного рефлекса световое раздражение, используемое при выработке оборонительного условного рефлекса, в ипсилатеральном полушарии не воспринимается как сигнальное. В этом, видимо, проявляются взаиморегулирующие отношения полушарий головного мозга при условно-рефлекторной деятельности.

Рис.7. Импульсная активность пары нейронов симметричных пунктов
зрительной коры.
Верхние нейронограммы — из ипсилатерального, нижние — из контралатерального относительно стороны подкрепления полушария. Цифры слева — номера сочетаний. Стрелка — момент предъявления сигнальной вспышки света. Отметка времени — 50 мс.
Однако кодирование информации не ограничивается характером импульсной последовательности [10]. Как показали наши исследования, на всех этапах условно-рефлекторной деятельности имеет место изменение взаимосвязи активности соседних нейронов в сторону ее усиления.
Анализ корреляционной зависимости импульсных потоков «обучающихся» и «необучающихся» нейронов показал, что значимые корреляционные отношения наиболее часто встречаются среди первых, т.е. при выработке рефлекса след об обучении сохраняется в рисунке взаимосвязи между нейронами, преимущественно участвующими в данном условно-рефлекторном процессе.
Указанные изменения нейронной активности в результате обучения были выражены в разных полушариях и на разных этапах упрочения условного рефлекса не однозначно.
В предсигнальный период на первом этапе выработки рефлекса, когда условной реакции у животных еще не было, не отмечалось и межполушарных различий в активности нейронов, хотя в фоне они имели место.
Общая реактивность исследуемых областей не отличалась и в период предъявления сигнала, но количество «обучающихся» нейронов было больше в контралатеральном относительно стороны подкрепления полушарии.
В период безусловной стимуляции в начале выработки рефлекса полушарных отличий активности нейронов зрительной коры также не выявилось. Это и понятно, так как еще не сформировалась условно-рефлекторная система, вызывающая асимметрию полушарного взаимодействия, и, как показано нами [58], индифферентный, неадекватный для зрительной коры стимул вызывает в этой зоне обоих полушарий одинаковое состояние активности.
В последействии на первом этапе выработки рефлекса особых различий в активности нейронов симметричных областей зрительной коры также не отмечено.
В соматосенсорной коре, как и в зрительной, на первом этапе уже были нейроны, реагировавшие на обучение, но по количеству «обучающихся» нейронов, по средней частоте их импульсов, вариабельности средней частоты, величине корреляции потоков импульсов межполушарных отличий не было, т.е. в этих зонах коры исчезали различия в активности полушарий, имеющие место в ответах на адекватную электрокожную стимуляцию до выработки рефлекса.
Следовательно, начало выработки рефлекса как бы стирает имеющиеся в фоне априорные различия нейронной активности зрительной и соматосенсорной коры полушарий.
На втором этапе, когда появляются первые односторонние условные реакции, картина отношений симметричных пунктов зрительной коры существенно меняется. В предсигнальный период возбудимость нейронов (о чем можно судить по средней частоте разрядов в единицу времени) стала выше в контралатеральном полушарии, здесь же стала больше и корреляция потоков импульсной активности одновременно зарегистрированных нейронов.
В период условной стимуляции в ипсилатеральном полушарии уменьшалось число нейронов, проявляющих признаки обучения, уменьшалось общее число нейронов, реагирующих на свет, но и среди последних увеличивалось количество тормозящихся. В контралатеральном полушарии реакции нейронов на сигнал были сильнее. Нужно отметить, что наряду с усилением корреляции между «обучающимися» нейронами, зарегистрированными внутри полушарий, увеличивалась корреляция между потоками активности нейронов, зарегистрированных в разных полушариях.
Для периода безусловной стимуляции было характерно проявление новых межполушарных отношений. Если ранее безусловный стимул, как неадекватный для зрительной коры, вызывал однотипное состояние ипсилатерального и контралатерального полушарий, то на втором этапе выработки рефлекса он чаще вызывал у нейронов контралатерального полушария возбуждение, а у нейронов ипсилатерального – торможение активности. Следовательно, с этого этапа в коре, организующей условную реакцию, начинают усиливаться тормозные процессы. Об изменении полушарного соотношения активности на втором этапе свидетельствует большое количество нейронов контралатерального полушария, сохраняющих в последействии следовые явления условной и безусловной стимуляции.
В соматосенсорной коре, как и в зрительной, в сигнальный период, в периоды подкрепления и последействия увеличивалась корреляция импульсных потоков «обучающихся» нейронов, число которых в разных полушариях было близким. Другие показатели нейронной активности изменялись незначительно. Однако, в предсигнальный период в контралатеральном полушарии относительное количество нейронов, изменяющих активность в ходе опыта, было выше, чем в ипсилатеральном.
На третьем этапе, когда имеет место билатеральная реакция на сигнал в обоих полушариях на всех стадиях рефлекса наблюдались однотипные процессы: снижение средней частоты разрядов, сокращение числа возбуждающихся и увеличение числа тормозящихся нейронов, уменьшение вариабельности частоты разрядов, усиление корреляционных отношений между потоками импульсов нейронов, проявивших признаки обучения.
В соматосенсорной коре на третьем этапе была достоверная межполушарная разница в количестве «обучающихся» нейронов – в контралатеральном полушарии их было больше в предсигнальный период, вариабельность их потоков импульсов ниже, чем у нейронов ипсилатерального полушария. Однако, несмотря на эти отличия, можно отметить, что на третьем этапе, как в зрительной, так и в соматосенсорной областях повышалось сходство состояния симметричных пунктов полушарий.
Дальнейшая выработка рефлекса приводила к усилению сходства полушарной активности зрительных зон коры. Так, на четвертом этапе, когда появились реципрокные двигательные реакции, в предсигнальный период, периоды подкрепления и последействия межполушарных отличий активности не отмечалось. Однако, на этом этапе во вторичной зрительной коре сигнальный период характеризовался сохранением большей частоты разрядов «обучающихся» нейронов в контралатеральном, а не в ипсилатеральном полушарии.
Отличия нейрональной активности разных полушарий четко сформировались на пятом этапе, т.е. при упроченном рефлексе. На этом этапе в зрительных и соматосенсорных областях коры число нейронов, изменяющих активность, «обучающихся», число пар нейронов со значимыми коэффициентами корреляции между потоками их импульсов было больше в контралатеральном относительно стороны подкрепления полушарии. Эти различия наиболее четко отмечались в период условной стимуляции.
Следовательно, как и в опытах с регистрацией суммарной, фокальной активности, так и опытах с регистрацией импульсной активности отмечалась единая динамика межполушарных отношений: в начале выработки рефлекса стирались существующие в фоне полушарные особенности активности коры, затем усиливалось сходство ее по всем исследуемым зонам, и, наконец, когда имела место локальная односторонняя двигательная реакция на сигнал, формировалась функциональная межполушарная асимметрия. Эта асимметрия не сопровождалась различиями средней частоты разрядов. И, если бы не разделение нейронов на «обучающиеся» и «необучающиеся», то межполушарных отличий выделить бы не удалось. Оказалось, что наибольшие различия между полушариями мозга заключались в количественном представительстве «обучающихся» нейронов, величине вариабельности средней частоты их разрядов, взаимоскоррелированности потоков их импульсных реакций. Активность этих нейронов, видимо, и создает в ответ на сигнал то состояние, которое необходимо для возникновения условно-рефлекторного ответа [47].
Установлена высокая корреляция отношений активности «обучающихся» нейронов, расположенных в коре одного и в коре разных полушарий, а также отсутствие подобных отношений у «необучающихся» нейронов, что соответствует представлениям М.М. Хананашвили и др. [97], В.Л. Силакова [85] о клеточных микросистемах, «предназначенных для реализации некоторой функции и выступающих по отношению к другим нейронам, объединенным как единое целое» [85]. Нейронные микросистемы образуются не только за счет нейронов корково-подкорковых структур одного полушария [42, 85 и др.], но и за счет тормозно-облегчающих влияний симметричных пунктов полушарий.
Следовательно, в процессе обучения формируется нейрональная многоуровневая микросистема, обеспечивающая условную реакцию. Естественным становится вопрос о нейрональном механизме формирования такой системы.
А.А. Ухтомский [93] отмечал, что в естественных условиях доминанта представляет собой разветвленную систему связей и возбуждений, объединенную какой-либо функциональной задачей. В наших исследованиях показано, что применение билатерального сигнального раздражения приводило к формированию в полушариях контрастных изменений возбудимости. В контралатеральном, относительно стороны подкрепляющего стимула полушарии в результате обучения увеличивается число «обучающихся» нейронов, которые обладают большей чувствительностью к сигнальным стимулам. Об этом говорит также возникновение на эти стимулы позднего позитивного потенциала в контралатеральном полушарии и исчезновении его в ипсилатеральном. Указанный феномен наблюдался на II-III-IV этапах образования условного рефлекса, но наиболее четко он имел место на V-м этапе, т.е. при упроченном рефлексе. Это может свидетельствовать о формировании стойкого очага повышенной возбудимости относительно сигнального стимула в контралатеральном полушарии.
Каким образом развивается такое доминирование. Судя по развитию тэта-подобных ритмов, ритма усвоения, позднего позитивного потенциала, паттерна импульсной активности нейронов в начале выработки рефлекса билатеральный сигнальный стимул вызывал увеличение различий в полушарной активности, что, видимо, объясняется усилением исходной доминантности, обеспечивающей большую активность ведущей лапы.
Под влиянием сочетаний наблюдалось параллельное развитие доминанты в обоих полушариях, что приводило к двусторонним двигательным эффектам. Затем наступал период конкурентной борьбы доминантных очагов. В этот период не происходило полного подавления активности одной из доминант, а, видимо, только временное смещение ее фокуса, что и выявлялось в форме реципрокных двигательных реакций.
Повторение сочетаний за счет одностороннего подкрепления при оборонительном условном рефлексе, или за счет односторонней инструментальной реакции при пищевом условном рефлексе, приводило к формированию доминантного очага, обеспечивающего условную реакцию в контралатеральном относительно стороны подкрепления. Эти данные можно расценить как то, что функциональная межполушарная асимметрия обучающегося мозга формируется на основе механизма доминанты. Видимо, постоянная афферентация к одним и тем же корковым проекционным системам за счет одностороннего подкрепления или за счет односторонней инструментальной реакции, в контралатеральном полушарии создается стационарное возбуждение, обеспечивающее посылку разряда в конечный путь, завершающийся конкретным рефлекторным актом. Такое проявление доминанты А.С. Батуев [11] определяет как вектор поведения организма при разнообразных влияниях окружающей среды.
Таким образом, на основании полученных в экспериментах результатов по электрической активности нейронов, фокальной активности и суммарной ЭкоГ, а также по результатам поведенческих реакций, прослеженным в динамике образования условного рефлекса, можно полагать, что в основе межполушарной асимметрии при реализации временной связи лежит формирование в полушарии доминантного очага, т.е. в основе природы функциональной межполушарной асимметрии и ее модуляции лежит доминантный механизм [16, 21].
Динамика межполушарных отношений не однозначна даже в области коры одного анализатора. Этот факт установлен нами при анализе нейронной активности первичных (17-е поле) и вторичных (18-е поле) зрительных полей.
Особенности нейрональной организации 17-х и 18-х зрительных полей выявились при анализе участия этих структур в динамике образования временной связи и на разных стадиях проявления условного рефлекса. Причем, эти особенности при качественной однозначности количественно отличались в ипсилатеральном и контралатеральном относительно стороны подкрепления полушариях (табл.1).
Приводимые разными авторами цифры о количестве нейронов зрительной коры, участвующих в условно-рефлекторной деятельности, весьма отличаются. Так, по данным Г.Н. Сметанкина [87], в этой коре на сигнал реагировало до 25% нейронов, по данным М.Я. Рабиновича [81] – 48%, а по данным А.П. Багдонас и др. [8] – около 6%. Видимо, такие различия обусловлены, прежде всего зоной исследования коры и этапом
выработки рефлекса, на котором была сделана выборка.
В наших исследованиях анализировались перестройки нейрональной активности в динамике выработки рефлекса в периоды: предсигнальный, сигнальный, подкрепления и после сочетания, а также определялась зависимость перестроек активности нейронов от их полушарной принадлежности.
Оказалось, что в первичных и вторичных зрительных полях обоих полушарий в предсигнальный период на двух первых этапах выработки рефлекса число нейронов, участвующих в организации условно-рефлекторной реакции, было близко. Начиная с третьего этапа и включая пятый, больше таких нейронов становится во вторичной зрительной коре (табл. 1).
Таблица 1
Число нейронов (в %) первичной (I) и вторичной (II) зон зрительной коры,
изменяющих активность в динамике выработки оборонительного условного рефлекса
|
Период анализа |
Этапы выработки рефлекса |
|||||||||
|
первый |
второй |
третий |
четвертый |
пятый |
||||||
|
I |
II |
I |
II |
I |
II |
I |
II |
I |
II |
|
|
Ипсилатеральное полушарие (относительно подкрепления) |
||||||||||
|
Предсигнальный |
57 |
60 |
60 |
54 |
55 |
71 |
30 |
55 |
3 |
7 |
|
Сигнальный |
51 |
63 |
76 |
76 |
67 |
77 |
44 |
77 |
55 |
58 |
|
Подкрепления |
33 |
66 |
52 |
60 |
58 |
17 |
28 |
15 |
9 |
16 |
|
Последействия |
24 |
37 |
27 |
45 |
16 |
27 |
28 |
12 |
9 |
13 |
|
% обучающихся ней-ронов (по сигналь-ному периоду) |
24 |
30 |
12 |
49 |
14 |
52 |
14 |
40 |
12 |
54 |
|
Контралатеральное полушарие (относительно подкрепления) |
||||||||||
|
Предсигнальный |
62 |
62 |
67 |
66 |
62 |
72 |
47 |
77 |
37 |
48 |
|
Сигнальный |
59 |
55 |
71 |
87 |
56 |
84 |
57 |
78 |
73 |
77 |
|
Подкрепления |
49 |
61 |
84 |
89 |
72 |
33 |
31 |
37 |
21 |
26 |
|
Последействия |
28 |
58 |
36 |
40 |
20 |
23 |
21 |
28 |
11 |
31 |
|
% обучающихся ней-ронов (по сигналь-ному периоду) |
34 |
39 |
24 |
54 |
24 |
53 |
22 |
60 |
18 |
67 |
В ипсилатеральном полушарии в период предъявления сигнала на первом этапе реагирующих нейронов было больше в 18-м поле, чем в 17-м; на втором этапе наблюдается уравнивание числа реагирующих нейронов в обоих полях; на последующих этапах – вновь увеличение в 18-м поле. В контралатеральном полушарии за исключением первого этапа выработки рефлекса количество реагирующих нейронов сравнительно с17-м полем было всегда больше в 18-м поле.
В период подкрепления в ипсилатеральном полушарии на первых двух этапах количество нейронов, реагирующих на подкрепляющий стимул, больше во вторичной коре, на третьем-четвертом – в первичной и, наконец, на пятом этапе превосходящее число таких нейронов вновь отмечалось во вторичной зрительной коре. Близкой к описанной была динамика количественных отношений нейронов контралатерального полушария за исключением четвертого этапа. Следует отметить, что в этом полушарии число реагирующих нейронов было больше, чем в ипсилатеральном.
В период последействия на большинстве этапов следовые процессы более выраженно сохранялись во вторичных зрительных полях контралатерального полушария.
Эти факты свидетельствуют о большей значимости в условно-рефлекторной деятельности 18-го поля зрительной коры, что подтверждается и преобладанием в нем, особенно в контралатеральном полушарии, числа «обучающихся» нейронов. Так, при упроченном рефлексе в 18-м поле ипсилатерального полушария признаки обучения отмечались у 54% нейронов, а в контралатеральном у 67%, в то время как в 17-м поле их было 12% и 18% соответственно.
Таким образом, нашими опытами установлено, что первичные зрительные поля также участвуют в условно-рефлекторной деятельности, но относительно незначительным количеством нейронов.
Большее участие вторичных (ассоциативных) зрительных полей обусловлено, по-видимому, их обширными морфологическими связями с симметричными областями полушарий [66, 12, 123].
Имея такие критерии условно-рефлекторных перестроек корковой активности, как проявление специфического узора в последовательности импульсной активности и возникновение в фокальном ответе на кондиционирующий стимул позднего позитивного потенциала, мы поставили задачу исследования характера активности симметричных зон коры в процессе обучения в условиях депрессии локального участка зрительной коры ипсилатерального относительно стороны подкрепления полушария. Оказалось, что в этих условиях скорость формирования специфического узора импульсной и фокальной активности в симметричном пункте контралатерального полушария была меньше, чем при интактных полушариях. Когда специфический узор импульсной и фокальной активности стабилизировался, то в обученном полушарии подвергали депрессии локальный участок, симметричный ранее выключавшемуся пункту ипсилатерального полушария. В этом случае в необученном пункте условно-рефлекторные перестройки активности формировались заново, но быстрее, чем в контралатеральном полушарии. Следовательно, недепрессированные области уже обученного полушария способствовали более быстрому возникновению условно-рефлекторных перестроек в участке коры, ранее не участвующего в этом процессе.
В другой серии экспериментов на фоне стабильно появляющегося в активном полушарии условно-рефлекторного ответа предъявлялся ряд сочетаний при совместной работе полушарий. Можно было ожидать, что при снятии депрессии специфический узор активности на сигнал возникнет сразу в ранее депрессированном участке и, конечно, сохраняется в этой зоне коры, где он уже был.
Однако этого не произошло. Как в ранее обученном полушарии, так и в необученном в начале совместной их работы в фокальном ответе отсутствовал поздний позитивный потенциал. Только повторение сочетаний вызывало постепенное восстановление характера фокальной активности в коре интактного полушария, а затем появление его в полушарии, которое ранее функционально выключалось. Статистическое сравнение величин ЛП и других характеристик нейрональной активности показало, что после снятия депрессии они становились сходными с таковыми до выработки рефлекса и только после предъявления сочетаний постепенно приобретали значения, присущие стадии упроченного рефлекса.
По данным Bures et al., [107], спонтанный межполушарный перенос обучения отсутствует, но его можно вызвать вставочными сочетаниями в промежутках между депрессиями. Билатеральный опыт облегчает выработку условного рефлекса в необученном полушарии. Bures et al., [107] считают, что вставочные тренировки вызывают перенос информации. Однако, по нашим данным, это не совсем так. Как показали опыты с фокальной и импульсной активностью, после снятия депрессии необученная зона коры мешает проявлениям условно-рефлекторной активности в ранее обученном симметричном пункте. Это проявляется в неспецифической генерализации активности. Следовательно, прямого переноса обучения не существует, а обучение происходит как бы заново. В этом случае генерализация исчезает намного быстрее, чем при первичном обучении.
Таким образом, в норме, когда функционируют оба полушария, специфическая активность при обучении возникает в обоих полушариях, хотя и с небольшим разрывом во времени. В основе этого процесса лежит механизм условно-рефлекторной генерализации.
Условно-рефлекторные эксперименты позволяют заключить, что у животных помимо врожденной межполушарной асимметрии функций, возникает функциональная асимметрия и в результате обучения. В этом плане мозг человека не является исключением. И, несмотря на то, что в литературе накоплен колоссальный материал, доказывающий наделенность каждого полушария человека способностями, слабо выраженными или отсутствующими в другом [113, 112, 111, 25], трудно согласиться с представлением о полной специализации полушарий головного мозга. Такое представление противоречит прежде всего самой сути основных свойств нервных структур мозга: их высокой пластичности, адаптивности, надежности, обучаемости. И действительно, любые нейронные системы левого и правого полушарий могут быть обучены реагировать на одни и те же сигналы. В наших опытах с выработкой оборонительного условного рефлекса, в частности, установлено, что латерализация функций в коре мозга зависит от условий эксперимента (например, от стороны мозга, в которую адресуется афферентация от подкрепления). К этому следует добавить, что в опытах с расщепленным мозгом показано, что правому полушарию, как и левому, присущи явления, которые относятся к логическому мышлению и пр. [116].
В связи с этим более импонирующим является представление о высококомпенсирующем, облегчающем сотрудничестве полушарий при организации и осуществлении той или иной нервно психической функции [53, 86, 72]. В этом плане наши эксперименты с выработкой оборонительного и пищевого условных рефлексов могут служить хорошей иллюстрацией. В них показано, что доминирование полушария в организации какой-либо функции может быть результатом обучения. Это же явление было отмечено и у человека на примере становления зрительно-моторной реакции в онтогенезе.
На основе опытов с регистрацией вызванных потенциалов В.Л. Бианки [18, 19, 23] выявлена парциальная асимметрия в зрительной системе полушарий. В то же время можно предположить, что такая асимметрия имеет место и в функционировании других анализаторов. Как показали морфологические и нейрофизиологические исследования, первичные и вторичные зоны коры одного анализатора связаны межполушарными комиссурами неодинаково. По-видимому, само разделение коры на первичные и вторичные области создает возможность парциального доминирования в пределах одного анализатора, так как трудно представить последнее в малодифференцированных структурах с эквипотенциальными функциями.
В наших исследованиях также выявлено наличие парциальной асимметрии у кошек. Оказалось, что не всегда животные, у которых доминировало левое полушарие (по более активной лапе), имели в 17-м и 18-м полях в этом полушарии и большее количество реагирующих нейронов. У некоторых животных с ведущей правой лапой количество реагирующих нейронов в зрительной коре было больше не в контралатеральном, а в ипсилатеральном полушарии.
W.G. Webster, [125] выявил факт корреляции предпочтения передней конечности с латерализацией анализа зрительных раздражений у кошек. Оказалось, что дифференцирование зрительных раздражений у части кошек происходили лучше в полушарии, ипсилатеральном предпочитаемой лапы. Но, все же, у большинства животных ведущая лапа определяла более активное полушарие.
Таким образом, парциальное доминирование у животных в ряде случаев проявляется несовпадением доминирующего по лапе полушария с более активным полушарием по реактивности нейронов зрительной коры. Кроме того, зрительная кора не целиком доминирует в одном полушарии. Об этом свидетельствует тот факт, что количество реагирующих элементов и другие показатели активности нейронов 18-го и 17-го полей одного полушария были не одинаковы.
Исходя из того, что в результате обучения, особенно при оборонительном условно-рефлекторном поведении, как правило, создаются условия преимущественной афферентации в одно полушарие, парциальное доминирование полушарий может быть обусловлено наряду с врожденными, и факторами внешних воздействий.
Исходная парциальная асимметрия 17-х и 18-х полей зрительной коры при выработке рефлекса усиливалась, если подкрепление адресовалось в доминирующее полушарие. Это сказывалось в изменении количества нейронов, участвующих в условно-рефлекторной деятельности.
При неадекватном для данной коры раздражении межполушарные отличия исчезают. Эти факты, по-видимому, означают, что в эволюции коры головного мозга помимо разделения ее областей на сенсорные зоны с последующей дифференцировкой в них первичных, вторичных зон, происходит дальнейшее их дифференцирование по уровню чувствительности к сенсорным раздражениям. Такое дифференцирование приводит к так называемой парциальной межполушарной асимметрии в пределах одного анализатора.
Что означает парциальное доминирование? Каков биологический смысл в этом процессе, возникшем в эволюции? В определенной мере ответ на эти вопросы получен в исследованиях В.М. Мосидзе и др. [71] и в нашей работе, показывающих, что при одностороннем доминировании полушария скорость выработки условных рефлексов, их переделка, скорость зрительно-моторной реакции, возбудимость нервных структур выше, чем при парциальном. Однако, последнее, проигрывая в скорости, выигрывает в устойчивости нервных процессов.
Рассматриваемые в данной работе формы условно-рефлекторного поведения, анализ фонового состояния коры полушарий и его изменений в условиях воздействий адекватных и неадекватных стимулов, а также в динамике образования временной связи, выявили ряд закономерностей в отношениях активности коры разных полушарий. Эти закономерности не являются порождением только внешних влияний на организм, но имеют и врожденную природу. О последнем свидетельствует то, что фоновая суммарная и импульсная активности, возбудимость коры в полушариях отличались неоднозначностью, которая, судя по изменению суммарной ЭкоГ, фокальным и импульсным ответам усиливалась при сенсорных воздействиях. Следует подчеркнуть, что существующие в фоне различия в активности полушарий усиливаются только при адекватной стимуляции (световой для зрительной коры, электрокожной для соматосенсорной и т.д.).
Оказалось, что выработка временной связи облегчается при подкреплении, не вызывающем противоречий с имеющейся асимметрией. В противном случае возникает необходимость преодоления фоновой асимметрии и создания ее вновь, но уже с доминированием другого полушария.
Это представление базируется на ряде исследованных нейрофизиологических показателей и на анализе поведенческих реакций в динамике выработки условных рефлексов разной формы – оборонительной и пищевой. Так, оказалось, что применение сочетаний условного и подкрепляющего сигналов в начале выработки рефлекса вызывает формирование однозначного состояния в симметричных пунктах разных полушарий. Чем она обусловлена?
По данным М.Н. Ливанова [64], John [116], Olds et al. [119], О.С. Адрианова [4]; В.Б. Швыркова [101] и др., при осуществлении поведения во многих структурах мозга происходит синхронизация активности. Однако, в этих работах не рассматривается вопрос о значении неадекватной стимуляции для синхронизации деятельности полей коры разных полушарий. Как нам представляется, исходно разная активность, возникающая в областях мозга, становится однотипной за счет сочетания двух разных по сенсорности стимулов, адекватных лишь для своей зоны коры, так как неадекватные стимулы не вызывают отличий активности между зонами коры полушарий. Следовательно, в динамике выработки рефлекса формирование однотипности импульсной, фокальной и суммарной активности между симметричными пунктами зрительной и соматосенсорной коры обусловлено (в наших опытах) сочетанием стимулов, один из которых является неадекватным для зрительной, другой – для соматосенсорной коры. Следовательно, сочетание неадекватной для зоны коры проекции сигнальной и подкрепляющей стимуляций является необходимым условием для синхронизации активности разных полушарий, а это в свою очередь, является одним из условий образования временной связи.
Существенным условием передачи возбуждения между корковыми зонами является пространственная синхронизация, в основе которой лежит q-ритм [91, 49]. Анализ механизма межполушарного переноса обучения (в наших экспериментах) позволил выявить, что при обучении одного полушария условно-рефлекторная активность в другом образуется лишь при совместной даже кратковременной их работе, которая приводит к однотипному состоянию симметричных пунктов коры. Последнее создает условия для межполушарного обмена условно-рефлекторной информацией. После его осуществления вновь формируется физиологическая межполушарная асимметрия характера активности.
Таким образом, полученные экспериментально материалы показали, что наблюдавшаяся в спокойном состоянии у животных относительная асимметрия биоэлектрических процессов симметричных пунктов коры разных полушарий в динамике выработки рефлекса под воздействием сочетания раздражителей меняется от усиления к стиранию различий и вновь к их усилению при упроченном рефлексе.
Такое развитие электрофизиологических коррелятов межполушарных отношений подтверждалось и в поведенческих опытах с произвольным выбором лапы для реакции на инструмент воздействия при выработке пищевого условного рефлекса.
Таким образом, в ходе формирования временной связи в условно-рефлекторный процесс постепенно вовлекаются оба полушария, а затем происходит концентрация возбуждения в одном из них. Какой в этом физиологический смысл? Не рациональней ли было бы при выработке рефлекса постепенно усиливать создаваемую односторонним подкреплением асимметрию, столь нужную для организации условно-рефлекторного ответа? Однако этого не происходит, условно-рефлекторная асимметрия формируется через стадию симметрии состояния. Возникновение при выработке рефлекса двух симметричных очагов возбуждения необходимо, так как они создают условия для фиксации в обоих полушариях однотипных энграмм памяти. Ввиду того, что контралатеральное полушарие, в которое афферентация от безусловного стимула поступает более интенсивно, занимает доминирующее положение, активность ипсилатерального постепенно тормозится. Однако, это торможение развивается лишь после того, как зафиксируется информация о значении сигнального стимула. Видимо, генерализация условного рефлекса на начальных этапах его выработки препятствует функционально доминирующему полушарию тормозить активность симметричных систем до тех пор, пока в их памяти не произойдет фиксации происходящих событий.
В настоящее время не вызывает сомнений факт функциональной специфичности полушарий головного мозга человека. Суммируя литературные данные можно выделить для каждого полушария преимущественно выполняемые ими функции.
Для правого полушария мозга характерно преимущество в процессах анализа невербальных сигналов: оно оценивает и воспроизводит положение объектов в пространстве, осуществляет пространственный анализ схемы тела и зрительных сцен лучше, чем левое полушарие. Люди с доминированием правого полушария отличаются медлительностью, в том числе речи, они предпочитают запоминать образный, а не логический материал, в своей деятельности опираться на личный опыт, а не переданный ему словесно, зрительная память его лучше, чем у левополушарного.
Левое полушарие лучше, чем правое анализирует вербальные сигналы, обеспечивает тонкие двигательные дифференцировки и жестикуляции правой руки. Левополушарные люди отличаются расторможенностью, эйфоричностью, проявляют более выраженные положительные реакции при восприятии вербальных сигналов, им свойственна легкость речи, они быстро запоминают словесный материал, легко используют чужой опыт, логическая память у них лучше, чем у правополушарных.
Кора левого полушария человека функционально более дифференцирована, чем правого [51]. Более активное доминирующее полушарие обладает большей частотой и меньшей амплитудой a -ритма [34, 36, 120, 105]. Внутриполушарные связи левого полушария короче, чем правого, что обусловливает фокальный тип активности в левом и диффузный в правом мозговом полушарии [22]. У правшей временные связи легче образуются в левом, а у левшей – в правом мозговом полушарии[61].
Принято считать, что формирование функциональной межполушарной асимметрии в онтогенезе является необходимым условием гармоничного развития личности.
Известно, что при старении изменения в организации зрительно-пространственного анализа не одинаковы в разных полушариях [78]. Наше исследование на разных возрастных группах показали, что межполушарная асимметрия в организации зрительно-моторной реакции активно развивается в онтогенезе к 6-7 годам жизни, хотя, как отмечает D.Taylor [124], впервые преимущественное использование одной руки у детей замечается уже к концу первого года. Однако, реакция активации ЭЭГ становится различной в левом и правом полушариях с 6-7 лет [94, 13, 38, 96].Существует значительное число исследований, посвященных вопросам периодизации в развитии ребенка, определения возрастных этапов, но эти работы в основном базируются на морфологических признаках (темпы роста, масса тела, смена зубов), социальных определениях (ясельный, дошкольный, младший школьный, старший школьный возраст). Наряду с этим, конечно, имеются разработки, посвященные изучению возрастной динамики психического статуса ребенка [95, 14, 37].
В литературе мало работ, которые учитывали бы роль парциального или одностороннего доминирования полушарий в созревании функциональной готовности различных систем организма. Наши исследования показали, что в случае парциального доминирования (более дробного распределения функций между полушариями, чем при одностороннем) созревание мозга происходит быстрее. Это, видимо, необходимо учитывать при определении оптимальных нагрузок на растущий организм [35, 5, 98]
А.Г. Хрипкова и др., [98] считают, что переломным этапом развития ребенка является возраст между 4 и 6-7 годами; к 6 годам происходят качественные перестройки процессов восприятия от генерализованных к локальным. К 6-7 годам происходит совершенствование точности движений, механизмов совершенствования произвольной регуляции двигательных актов [30]. Возраст 6-7 лет является критическим этапом установления новых взаимоотношений эндокринной системы [83, 52].
Исходя из сказанного, наши данные о различии зрительно-моторной реакции при парциальной асимметрии с 6 лет, а при односторонней асимметрии с 7 лет, получают определенную обоснованность.
Исходя из собственных наблюдений и литературных данных [68], нами высказано предположение, что асимметрия полушарий головного мозга в организации зрительно-моторной реакции формируется в онтогенезе как результат обучения. Это предположение основывается, например, на том, что распределение латентных периодов реакций левой и правой руки у детей 4-5 лет имело мономодальный характер. У детей с парциальным доминированием, начиная с 6-летнего возраста, а у детей с односторонним – с 7-ми летнего, в таком распределении ЛП реакций ведущей руки появляются два пика: относительно коротколатентный и длиннолатентный. В то же время ЛП реакций субдоминантной руки имели один пик в области длиннолатентных величин (рис. 8).

Рис. 8. Нормированные гистограммы распределения ЛП зрительно-моторных реакций испытуемых с парциальным (вверху) и односторонним (внизу) доминированием мозга. Цифры слева соответствуют возрастным группам: 1 – 4 года, 2 –5 лет, 3 –6 лет, 4 – 7 лет, 5 –15 лет, 6 – 18-20 лет, 7 – 21-30 лет.

Само по себе это различие еще не говорит о том, что межполушарная асимметрия формируется как результат обучения. Однако с возрастом формирование двух пиков в распределении ЛП реакций ведущей руки становится более четким. Следовательно, тренировка, частое использование одной руки в двигательных актах способствует появлению более коротких ЛП. Второй факт, который также подтверждает роль обучения в функциональной асимметрии полушарий, заключается в том, что коротколатентные ответы исчезают у испытуемых с развитием утомления (рис.9).


Рис.9. Нормированные гистограммы распределения ЛП зрительно-моторных реакций. Вверху: у испытуемых рабочих в начале и в конце смены. Внизу: у школьников в начале и конце полугодия. О – испытуемые с односторонним, П – парциальным доминированием мозга.
Известно, что условно-рефлекторная часть поведения более утомляема, чем, безусловно-рефлекторная, и что левое полушарие у человека преимущественно связано с процессами обучения. Отсюда появление коротких ЛП ответов, организуемых левым полушарием и исчезающих при утомлении, можно считать результатом обучения.
То, что межполушарная асимметрия обеспечивает большую быстроту выполнения зрительно-моторной реакции доминирующей руки, не означает, естественно, что этим исчерпывается ее функциональное значение. Еще одно из ее значений было выявлено при анализе ЭЭГ-активности симметричных систем мозга в динамике развития утомления.
Оказалось, что в обоих полушариях с ухудшением работоспособности испытуемых увеличивается a-индекс, удлиняются периоды a-волн, снижается возбудимость коры, межполушарная асинхронность колебаний a-ритма либо значительно ослабевала, либо чрезмерно усиливалась. Высокая работоспособность, как правило, коррелировала с определенным, по-видимому, оптимальным для данного индивида уровнем межполушарной асинхронности.
В литературе [46, 44, 118] показано, что при утомлении усиливается межполушарная асимметрия активности.
В наших исследованиях утомление сопровождалось изменением уровня межполушарной асинхронности активности. Однако это изменение могло быть двунаправленным – в сторону усиления или в сторону ослабления. Противоречие в этом случае решается, видимо, тем, что для оптимальной работоспособности нужен и оптимальный уровень асимметрии идентичных систем разных полушарий. И действительно, снижение асимметрии или ее резкое увеличение приводят к росту относительного количества ошибок. Для нормальной работоспособности человека необходим определенный для данного индивида уровень асинхронности, а, следовательно, определенный уровень функциональной межполушарной асимметрии в активности симметричных пунктов полушарий головного мозга. Превышение или снижение этого уровня нарушает работоспособность.
Развитие исследований межполушарного взаимодействия и распределения функций между полушариями головного мозга показало, что доминирование левого полушария у человека не является полным. Так, например, пространственный анализ зрительных сигналов обеспечивается в основном корой правого полушария. Выявлено множество функций, преимущественно организуемых либо левым, либо правым полушарием [115, 117, 121]. Эти факты позволили сформулировать представление о парциальном доминировании, т.е. превалировании полушария в организации одной функции и недоминировании его в организации другой.
И у детей и у взрослых межполушарная асимметрия в покое выражена слабее, чем при умственной деятельности. У здорового взрослого человека в состоянии покоя межполушарная асимметрия сглаживается, сигнал к работе и любая ориентировочная реакция увеличивают ее, а монотонная работа – уменьшает. При решении арифметических задач амплитуда колебаний ЭЭГ-активности снижалась в левом полушарии, оставаясь без изменений в правом [108]. Предъявление вербальной информации вызывают более выраженную депрессию a -ритма в левом полушарии мозга. Характер межполушарного взаимодействия зависит от специфики задачи: при решении логических задач фокус изменения a -активности у правшей отмечается в левой лобно-височной области, а при слежении за световым сигналом – в симметричных затылочных областях [84].
Исследования показывают, что у детей правое полушарие более активно, чем у взрослых, участвует во всех видах умственной деятельности [50, 94].
Проблема парной деятельности мозговых полушарий интересна также с точки зрения их взаимодействия при разных функциональных состояниях человека и на различных фазах работоспособности. Электрографические исследования Л.П Павловой и др., [75] свидетельствуют о различной динамике межполушарных отношений в процессе развития утомления. По данным Г.А. Сергеева и сотрудников [84] при развитии умственного утомления фокус максимальной активности смещается в правое полушарие мозга. Аналогичные данные приводятся в работе Е.Б. Сологуб [88] при мышечных нагрузках: при врабатывании и утомлении «меченные» ритмы регистрируются в правом, а в период высокой работоспособности – в левом полушарии мозга.
У лиц с высоким уровнем интеллекта средняя частота ЭЭГ в лобных и теменных областях выше, чем у людей с обычным уровнем интеллекта.
Таким образом, в настоящее время исследованы некоторые электрографические проявления межполушарной асимметрии у человека, однако вопросы, связанные с ее значением при разных формах деятельности человека, в процессах связанных с обучением, а также вопросы развития асимметрии в онтогенезе остаются далеко не решенными.
Несмотря на факты, свидетельствующие о существовании различий в функциональной организации полушарий [26, 57, 69] в литературе встречается мало работ, в которых исследуются особенности пространственно-временной организации ЭЭГ-активности в зависимости от профиля латеральной организации (ПЛО) мозга человека. В литературе принято выделять 3 основных вида ПЛО: правый, левый и смешанный [26].
В нашей работе исследовались характеристики a -ритма симметричных областей мозга в динамике зрительного сканирования у правшей с ведущим правым (одностороннее доминирование) и левым (парциальное или смешанный тип доминирования) глазом. Обнаружено, что у большинства правоглазых правшей (ПП) альфа индекс в исходном фоновом состоянии был выше в затылочных и двигательных зонах коры правого полушария мозга. В процессе развития утомления отмечалось возрастание a -индекса в левом полушарии мозга и изменение асимметрии a -волн, свидетельствующее о снижении уровня возбудимости в обоих полушариях мозга. Параллельно отмечалась синхронизация активности симметричных зон коры и снижение эффективности зрительного сканирования. Причем периоду необнаружения сигнала соответствовало ослабление возбудительного процесса в коре.
Таким образом, асинхронность в активности мозговых полушарий является необходимым условием высокой работоспособности человека.
У правшей с ведущим левым глазом (парциальное доминирование) величины a -индекса в обоих полушариях в исходном состоянии были близкими и недостоверно изменялись в динамике зрительного сканирования. Показатели возбудимости и асинхронность колебаний ЭЭГ в группе с парциальным доминированием была ниже, чем в группе с односторонним мозговым доминированием.
Исследование показателей спектральной мощности фоновой ЭЭГ в состоянии спокойного бодрствования с закрытыми глазами у испытуемых с разным ПЛО показало, что межгрупповые различия спектров мощности ЭЭГ отмечаются в левом полушарии мозга. Так у испытуемых с парциальным доминированием мощность D-активности в левой лобной коре (F3) в два раза выше, чем у лиц с односторонним левополушарным типом доминирования (праволатеральным ПЛО). Статистически достоверные различия спектральной мощности ЭЭГ отмечались и в альфа диапазоне: средняя мощность a -ритма в левом теменном отведении была ниже у «левополушарных» испытуемых по сравнению с этим же показателе у испытуемых с парциальным доминированием. В спокойном состоянии отмечались также различия пространственных характеристик ЭЭГ Внутриполушарная когерентность была выше у испытуемых с парциальным доминированием, а межполушарная – у лиц с односторонним левополушарным доминированием (рис. 10).

Рис.10. Усредненные значения функции когерентности фоновой ЭЭГ испытуемых с односторонним (толстая линия) и парциальным (тонкая линия) доминированием мозга. Статически значимые различия отмечены звездочками.
Таким образом, в состоянии спокойного бодрствования отмечается более высокий уровень активации левого полушария мозга испытуемых с праволатеральным ПЛО по сравнению с лицами со смешанным ПЛО. У последних за счет более мощных внутриполушарных связей полушария функционируют независимо друг от друга, в то время как у праволатеральных испытуемых межполушарное взаимодействие протекает на более высоком уровне, что может способствовать их объединению в единую интегрированную систему.
Динамика ЭЭГ показателей симметричных зон коры головного мозга человека в процессе интеллектуальной деятельности.
Исследовались показатели ЭЭГ и усредненных зрительных ВП (УЗВП) в процессе зрительного распознавания вербальной (слова с пропущенными буквами) и образной (невербализуемые фигуры А.Р. Лурия) у испытуемых с односторонним и парциальным доминированием мозга. Вербальный и пространственный тесты предъявлялись испытуемым на карточках на фоне световых вспышек. УЗВП служили индикатором уровня активации мозга (по показателям их блокирования в процессе вербальной и образной интеллектуальной деятельности по сравнению с состоянием спокойного бодрствования с открытыми глазами). В исследовании принимали участие 22 студента Ростовского государственного университета в возрасте от 20 лет.
Предварительно все испытуемые проходили психометрическое обследование, включающее регистрацию времени простой и сложной реакции на зрительные и слуховые раздражители, критической частоты слияния мельканий (КЧСМ), частоты теппинга правой и левой рукой.
Анализ данных психометрического обследования выявил преимущество в скорости сенсомоторных реакций и показателей подвижности структур зрительной и двигательной сенсорных систем у испытуемых с односторонним левополушарным типом доминирования мозга по сравнению с лицами с парциальным мозговым доминированием.
Сравнение амплитудно-временных характеристик УЗВП показало, что у испытуемых с парциальным доминированием в процессе интеллектуальной деятельности УЗВП блокируются в обоих полушариях мозга независимо от характера деятельности (вербальная или образная). У лиц с односторонним левополушарным доминированием мозга в процессе интеллектуальной деятельности УЗВП блокировались в меньшей степени, чем у испытуемых с парциальным доминированием, причем блокада УЗВП была более выражена преимущественно в левом полушарии мозга, что указывает на его преимущественную активацию при зрительном распознавании вербальной и образной информации. Качество деятельности, оцениваемое по количеству ошибок, было выше у испытуемых с односторонним доминированием, чем у лиц с парциальным доминированием.
То есть, одностороннее левополушарное доминирование мозга обеспечивает более высокую скорость и безошибочность интеллектуальной деятельности испытуемых. Это преимущество, по-видимому, базируется на формировании асимметричного состояния мозговой активности (левополушарной активации) в процессе зрительного распознавания слов и фигур. Симметричная активация мозговых полушарий испытуемых с парциальным доминированием, вероятно, оказывает отрицательное влияние на скорость и качество деятельности испытуемых.
Анализ спектральных и пространственных характеристик ЭЭГ симметричных лобных (F3,F4), центральных (C3,C4), теменных (P3, P4), затылочных (O1, O2) и височных (T3,T4) отведений показал, что у “левополушарных” испытуемых с правым профилем ПЛО при распознавании слов активируются преимущественно лобная кора левого полушария, а у испытуемых с парциальным доминированием – его затылочные области, а активация лобных зон коры осуществляется симметрично.
У лиц с правым ПЛО при распознавании слов отмечался низкий уровень когерентности как внутри- так и межполушарной (рис.11).

Рис. 11. Процентное соотношение высоких, средних и низких значений функции внутри- и межполушарной когерентности у испытуемых с односторонним левополушарным (правый профиль ФМА) и парциальным (смешанный профиль ФМА) типами доминирования мозга.
У испытуемых с парциальным доминированием мозга в процессе вербальной деятельности отмечался высокий уровень внутриполушарной когерентности при среднем уровне межполушарной когерентности биопотенциалов морга. Последнее, по данным литературы [31], может служить указанием на параллельное участие мозговых полушарий в процессах вербальной деятельности у испытуемых с парциальным доминированием мозга.
Наиболее высокое качество гностической деятельности при распознавании невербализуемых фигур отмечалось у испытуемых со смешанным типом мозгового доминирования. Качество распознавания невербализуемых геометрических фигур испытуемых с праволатеральным ПЛО было достоверно ниже, чем у лиц с парциальным доминированием. Испытуемые с односторонним правополушарным доминированием обнаруживали наихудшие показатели при распознавании невербализуемых фигур (рис.12).
Рис.12. Усредненные значения функции когерентности ЭЭГ для испытуемых с односторонним и парциальным типами мозгового доминирования. Различия статистически значимы для отведений CS-FS,OS-CS, OS-FS, O1-O2, OD-FD, OD-CD, CD-FD.
Анализ спектров мощности ЭЭГ при эффективном и неэффективном зрительном распознавании выявил достоверные различия спектральных характеристик ЭЭГ в ситуации успешного и неуспешного зрительного распознавания у всех обследованных испытуемых независимо от их ПЛО. Эти различия проявлялись в изменении ритмических составляющих ЭЭГ. В ситуации успешного распознавания предъявляемых фигур спектры ЭЭГ были более сглажены. В ситуации, когда опознать эталонную фигуру среди тестовых испытуемому не удавалось, в спектрах ЭЭГ появлялись дополнительные пики в диапазонах q -, a -, а иногда и b - частот. (рисунок 13).

Рис. 13. Спектры мощности ЭЭГ правой затылочной области коры при эффективном и неэффективном зрительном распознавании.
Пояснения в тексте.
Полученные результаты свидетельствуют о том, что в ситуации неуспешного зрительного распознавания предъявляемых зрительных стимулов имело место снижение уровня активации коры больших полушарий по сравнению с ситуацией эффективного распознавания фигур. Электрографические показатели в этом состоянии отражали усиление активности синхронизирующих систем мозга, что проявляется в появлении дополнительных пиков в спектрах мощности ЭЭГ преимущественно в диапазонах a - и q - частот.
Основные различия пространственно-временной организации ЭЭГ при зрительном распознавании геометрических фигур заключались в более высоком уровне психоэмоционального напряжения в процессе зрительного распознавания образной информации: в лобных зонах коры правого полушария мощность медленных ритмов была выше у лиц со смешанным ПЛО мозга по сравнению с испытуемыми с правым ПЛО как при успешном, так и при неуспешном зрительном распознавании, а также более высоких значениях внутриполушарной когерентности во время распознавания зрительной информации у лиц со смешанным типом мозгового доминирования по сравнению с односторонним левополушарным доминированием мозга (рисунок 4).
Поскольку правые лобные зоны коры относят к зонам, связанным с генерацией эмоций и, в частности, отрицательных эмоций, то наблюдаемые различия могут, с нашей точки зрения, отражать различия эмоционального статуса испытуемых с разным ПЛО мозга. Повышенный эмоциональный статус испытуемых со смешанным ПЛО находит свое отражение в более высокой активации у них эмоциогенных структур, в первую очередь структур лимбической системы, что выражается в повышенной мощности у этих испытуемых медленноволновой части спектра по сравнению с "левополушарными" испытуемыми с правым ПЛО. При распознавании невербализуемых фигур более высокие значения внутриполушарной когерентности отмечаются у лиц со смешанным доминированием полушарий по сравнению с односторонним как в передних, так и задних отделах левого полушария мозга.
Межполушарная когерентность, напротив, более высокая у лиц с левополушарным доминированием в зрительных областях коры, что может указывать на более тесное взаимодействие у них этих областей по сравнению с испытуемыми со смешанным типом мозгового доминирования. По-видимому, эти различия обусловливают индивидуальную стратегию зрительного опознания у лиц с разным типом мозгового доминирования, и лежат в основе наблюдаемых различий качества их деятельности.
Значение профиля латеральной организации мозга в процессах адаптации
Индивидуальные особенности функциональной межполушарной асимметрии мозга человека (ПЛО) тесно связаны с процессами адаптации организма в системе человек - среда. Показано, что адаптация к физическим и психическим стрессирующим факторам, тип вегетативной регуляции, «физиологическая цена» интеллектуальной деятельности, уровень работоспособности, утомления различны у лиц с разным профилем асимметрии мозга [7, 26, 40, 58, 59].
Освоение и использование спортивных двигательных действий, а также эффект специальной тренировки, возникающий у человека в процессе спортивной деятельности, обусловлены особенностями его морфо-функциональной организации, среди которой одно из главных мест принадлежит билатеральной асимметричности двигательного аппарата. Теоретический и практический аспект использования знаний об асимметриях функции парных органов человека в спорте наиболее полно обсуждается в исследованиях В.М. Лебедева [88], подчеркивающего необходимость их учета в «решении вопросов физиологии спорта. Возможность такого подхода диктуется самой природой человеческого организма, а необходимость - еще далеко не выясненным значением этого явления в практике спорта». Данные литературы по этому вопросу свидетельствуют, что моторная асимметрия, с одной стороны, связана со спецификой конкретного вида спорта, а с другой стороны, ее динамика отчетливо проявляется в зависимости от уровня подготовленности, стажа занятий и других факторов [39].
Исследовалась взаимосвязь индивидуальных особенностей межполушарной асимметрии мозга человека (ПЛО) и способности организма адаптироваться к физическим нагрузкам различного уровня по показателям регуляции сердечного ритма. В исследовании принимали участие студенты, регулярно не занимающиеся спортом в количестве 30 человек и студенты института физкультуры - 30 человек, в числе последних группа спортсменов высшей квалификации - мастеров спорта международного класса (10 человек).
Анализ ПЛО у обследуемых групп испытуемых: студентов РГУ, студентов ИФК и мастеров спорта показал, что по таким показателям зрительно-моторной асимметрии как ведущая рука, ведущее ухо и ведущий глаз, наиболее выраженной правосторонней латерализацией обладали мастера спорта, а наименее выраженной - студенты ИФК. Среднее положение по этим показателям занимали студенты-биологи, регулярно не занимающиеся спортом. Исследование особенностей ФМА мозга студентов, регулярно не занимающихся спортом, физкультурников и мастеров спорта показало, что большинство обследованных студентов имели правый или преимущественно правый профиль функциональной межполушарной асимметрии. У мастеров спорта зрительно-слухо-моторная асимметрия была наиболее высокой. Студенты ИФК имели минимальную степень латерализации функций по сравнению с другими обследуемыми.
Исследование показателей регуляции сердечного ритма у этих групп студентов показало, что доля нормальных показателей суммарного эффекта регуляции (нормосистолия) была максимальной у студентов ИФК и минимальной у мастеров спорта. Функция автоматизма сердца также была максимально близка к норме у студентов ИФК. Среди студентов, не занимающихся спортом, почти у половины испытуемых этот показатель отличался от нормы (чаще всего для них была характерна тахикардия). У мастеров спорта в 50% случаев отмечалась брадикардия, что типично при больших физических нагрузках. По показателю устойчивости регуляции сердечного ритма лидировали мастера спорта. По-видимому, увеличение этого показателя у мастеров спорта отражает долгосрочные адаптивные сдвиги в организме при усиленных тренировках.
Таким образом, полученные результаты свидетельствуют о том, что оптимальное состояние регуляции ритма сердца связано с умеренными физическими нагрузками. Интенсивные физические нагрузки у мастеров спорта вызывают развитие одних механизмов регуляции за счет других. Отсутствие физических нагрузок приводит к выраженным нарушениям регуляции ритма сердца в виде тахикардии в состоянии покоя, а также наличии дисрегуляции центрального типа, что характерно для студентов, подверженных значительным интеллектуальным нагрузкам и не занимающихся спортом. У студентов ИФК и мастеров спорта в результате адаптации к физическим нагрузкам развивается брадикардия и умеренная синусовая аритмия. Эти изменения могут отражать динамику адаптационных процессов в ходе тренировок и зависят от их интенсивности.
Важным показателем регуляции сердечного ритма является баланс вегетативных систем и связанный с ним вегетативный тонус. Анализ полученных результатов показал, что для большинства студентов ИФК была характерна нормотония, т.е. сбалансированность систем, управляющих работой сердца. Минимальное количество нормотоников отмечалось среди мастеров спорта (юношей и девушек). Студенты РГУ занимали по этому показателю среднее положение, среди них преобладали симпатотоники. В группе мастеров спорта отмечалась тенденция к преобладанию ваготоников, что может быть связано с долговременными адаптационными сдвигами в организме при больших и систематических физических нагрузках. Необходимо отметить, что вегетативный тонус у девушек был ниже, чем у юношей во всех обследуемых группах, что может быть связано с половыми различиями гормонального статуса испытуемых.
Другим важным показателем адаптационных возможностей и резервов организма является его реакция на нагрузку, которая является важным показателем состояния регуляторных систем организма в целом, а также их резервов и функциональных возможностей. В нашей работе в качестве нагрузки использовалась ортостатическая проба (переход из положения «лежа» в положение «стоя»).
Доля испытуемых с нормальной реакцией на нагрузку была наиболее высокой среди мастеров спорта (юношей и девушек) и студентов ИФК. Среди студентов, не занимающихся спортом, только половина девушек и 40% юношей обладали нормальной реактивностью, а у остальных реакция на ортостаз была либо избыточной, либо недостаточной. Причем среди студентов, не занимающихся спортом, в целом преобладали лица с повышенной реакцией на нагрузку, что может быть связано с недостаточной тренированностью организма, ограниченностью его функциональных резервов.
Таким образом, проведенное исследование показало, что оптимальное состояние организма обследуемых и их адаптационных резервов и возможностей коррелирует со смешанным типом доминирования мозговых полушарий. При больших физических нагрузках усиливается латерализация одних систем за счет других. Отсутствие физических нагрузок при интенсивной умственной деятельности приводит к усилению правосторонней асимметрии и активации симпатического и центрального контуров регуляции ритма сердца, что снижает функциональные резервы и возможности организма.
Список литературы
Абрамян Л.А. Принцип симметрии и диалектика. // Известия АН АрмССР, 1961, №1. С. 3-12.
Абуладзе К.С. К вопросу о функции парных органов. // М.: Медгиз, 1961. 104с.
Адрианов О.С. Об особенностях структурной организации функциональной межполушарной асимметрии мозга. // Взаимоотношения полушарий мозга: Материалы всесоюзн. конф. Тбилиси, 1982. С.147.
Адрианов О.С. О принципах организации интегративной деятельности мозга. М.: Медицина. 1976. 176с.
Алферова В.В., Горев А.С., Дубровинская Н.В. Возрастные особенности и нейрофизиологические механизмы произвольной регуляции внимания. // Новые исследования по возрастной физиологии. М.: Изд. АПН СССР. 1982, №1 (14). С.7-10.
Ананьев Б.Г. Билатеральное регулирование как механизм поведения. // Вопросы психологии. 1963, №5. С.83-98.
Аршавский В.В. Межполушарная асимметрия в системе поисковой активности. // Автореф. дисс. докт. биол. наук. Л., 1990. 41с.
Багдонас А.П., Полянский В.Б., Соколов Е.Н. Участие нейронов зрительной коры кролика в условном рефлексе на время. // Ж. высш. нервн. деят. 1968, т.18, №5. С.791-798.
Балонов Л.Я., Деглин В.Я. Слух и речь доминантного и недоминантного полушарий. // Л.: Наука. 1976. 218с.
Батуев А.С., Куликов Г.А., Каменская В.Г., Футер Л.И. кодирование биологической значимости звукового сигнала в коре мозга кошки. // Проблемы нейрокибернетики. Нейрофизиологические аспекты. Ростов-на-Дону. 1978. С.13-15.
Батуев А.С. Принципы конвергенции и доминанты. // Физиол. ж. СССР. 1976, т.62, №6. С.817-824.
Батуев А.С., Верещагина Е.М., Пирогов А.А. Особенности функциональных взаимоотношений парных отделов специфических и ассоциативных систем мозга животных. // Вестник Ленинградского ун-та. 1975, №3. С.60-67.
Бетелева Т.Г. Участие проекционной и непроекционной областей коры в сенсорном анализе у детей 3-10 лет. // Физиология человека. 1980, т.6, №5. С. 776-783.
Бетелева Т.Г., Дубровинская Н.В., Фарбер Д.А. Сенсорные механизмы развивающегося мозга. // М.: Наука. 1977. 176с.
Бехтерева Н.П. Нейрофизиологические аспекты психической деятельности человека. //Л.: Медицина. 1974. 151с.
Бианки В.Л. Эволюция парной функции мозговых полушарий. // Л.: Изд. Ленинградского университета, 1967. 260с.
Бианки В.Л., Удалова Г.П., Михеев В.В. Полушарная специализация у крыс при дифференцировании зрительных стимулов. // Ж. высш. нервн. деят., 1981, т.31, вып.1. С. 180-182.
Бианки В.Л. Индивидуальная и видовая межполушарная асимметрия у животных. // Ж. высш. нервн. деят., 1979, т.29, №2. С. 295-303.
Бианки В.Л., Абдуахадов А.С. Взаимодействие симметричных кортикальных доминант, сформированных при одновременной двусторонней поляризации. // Ж. высш. нервн. деят., 1980, т.30, №1. С.88-96.
Бианки В.Л., Курочкин В.А. Анализ межполушарных взаимоотношений в зрительной коре кошек на основании закона контрастного восприятия света. // Ж. высш. нервн. деят. 1975, т.25, №6. С.1061-1067.
Бианки В.Л., Филиппова Е.Б. Бинокулярное взаимодействие в парастриарной коре крыс при изменении интенсивности одного из монокулярных стимулов. // Нейрофизиология. 1973, т.5, №2. С.128-132.
Бианки В.Л. Механизмы парного мозга. // Л.: Наука. 1989. 264с.
Бианки В.Л., Харитонов Е.В., Шрамм В.А. Асимметричность фокуса максимальной активности транскаллозальных ответов. // Физиол. ж. СССР. 1978, т.64, №7. С.897-904.
Брагин А.М. Влияние одностороннего выключения коры больших полушарий головного мозга кошек на тонические и фазные инструментальные рефлексы в условиях переключения. // Взаимоотношения полушарий мозга: Мат-лы Всесоюзной конференции. Тбилиси, 1982. С.15.
Брагина Н.Н., Доброхотова Т.А. Функциональные асимметрии человека. // М.: Медицина, 1981. 288с.
Брагина Н.Н., Доброхотова Т.А. Функциональные асимметрии человека. // М.: Медицина, 1988, 240 с.
Бусель Б.И. Реакции нейронов зоны СI при оборонительном условном рефлексе на звук. // Нейрофизиология. 1975, т.7, №2. С.126-133.
Вайткявичус Г.Г., Соколов Е.Н., Фомин С.В., Блознялис М.О. О принципах максимальной чувствительности в системах детекторов. // Биофизика. 1974, т.19. С.154-161.
Василевский Н.Н. Нейрональные механизмы коры больших полушарий. // Л.: Медицина. 1968. 191с.
Васильева О.Н. Совершенствование физиологического механизма управления точностными движениями у детей. // Физиология развития человека. Тезисы II Всесоюзн. конф. М. 1981. С.214.
Вольф Н.В. Функциональная организация процессов полушарной обработки речевой информации. // Автореф. докт. дисс., Новосибирск, 1997. 34с.
Генкин А.А. Об асимметрии длительности возрастающих и убывающих фаз электроэнцефалограмм задних отделов головного мозга здорового человека. // Докл. Акад. пед. наук РСФСР, 1962, №4. С.99-102.
Глезер В.Д., Дудкин К.Н., Иванов В.А., Кульков А.П. О кодировании сигналов нейронами зрительной коры. // Исследование принципов переработки информации в зрительной системе. Л.: Наука. 1970. С.86-94.
Голиков Н.В. Электрическая активность головного мозга и ее зависимость от функционального состояния нервной системы. // Уч. зап. ЛГУ. Серия биол. наук. 1950. Т.22. С.202-228.
Громбах С.М. Принципы возрастной периодизации в гигиене детей и подростков. // Биологический возраст и возрастная периодизация. М.: 1978. С.5
Дарвей Б., Смык К. Межполушарная асимметрия амплитуды – ритма в электроэнцефалографической записи и проблема доминантности полушарий мозга. // Вопросы психологии. 1972, №3. С. 149-154.
Денисова З.В., Брендстед А.Н., Тараканов П.В. Межполушарная асимметрия тета-активности у детей при эмоциональных нагрузках. // Ж. высш. нервн. деят. 1978, т.28, №1. С.169-171.
Дубровинская Н.В. Формирование процессов активации коры головного мозга у детей по данным ЭЭГ. // Физиология человека. 1983, т.9, №1. С.33-42.
Ермаков П.Н. Психомоторная активность и функциональная асимметрия мозга. // М.: Изд. МГУ. 1988. 166с.
Ефимова И.Е., Хомская Г.Д. Межполушарная асимметрия функций и вегетативная регуляция при интеллектуальной деятельности // Физиология человека. 1990, №5. С.147-154.
Заркешев Э.Г. Нейронные механизмы корковой интеграции. // Л.: Наука. 1980. 144с.
Заркешев Э.Г. Нейронные механизмы корковой интеграции. // Автореф. дисс. докт. мед. наук. Ленинград. 1982. 40с.
Иваницкий А.М. Корковый синтез и две информационные системы мозга. // Ж. высш. нервн. деят., 1973, т.23, №2. С.375-383.
Караджов К.В. Исследование электрофизиологических показателей при различных видах умственной деятельности и развитии утомления. // Автореф. дисс. канд. мед. наук. М. 1976. 24с.
Карпинская Р.С. Идея сохранения и принцип симметрии в современной биологии. // Принцип симметрии. Историко-методологические проблемы. М.: Наука, 1978. С.303-318.
Касьянов В.М., Хан А.Б. влияние учебных занятий на согласованность в работе больших полушарий головного мозга школьника. // Детский организм и среда. М. 1973. С.19-33.
Книпст И.Н., Курова Н.С., Кориневский А.В. Динамика топограмм потенциалов и функциональное состояние коры больших полушарий. // М.: Наука. 1982. 166с.
Книпст И.Н. Биоэлектрическая активность разных уровней коры головного мозга при выработке оборонительного условного рефлекса на неритмические раздражения у кроликов. // Труды Института ВНД. Серия физиол. 1958. т.3. С.59-67.
Книпст И.Н., Кориневский А.Б., Курова Н.С. Динамика пространственных соотношений биопотенциалов коры больших полушарий. // М.: Наука. 1976. 248с.
Князева М.Г. Межполушарная асимметрия альфа-ритма ЭЭГ в процессе когнитивной деятельности различной успешности. // Физиология человека, 1991, т.17, вып.5. С.50-60.
Кок Е.П. (1967) Общее и различное в высших функциях симметричных отделов правого и левого полушарий мозга. // Физиология человека. 1975, т.1, №3. С.427-439.
Колесов Д.В., Чемоданов В.И., Сельверова Н.Б. Система гипофиз-гонады в период полового созревания у мальчиков. // Новые исследования в возрастной физиологии. 1980, №1. С.72.
Костандов Э.А. Функциональная асимметрия полушарий и эмоции. // Взаимоотношения полушарий мозга: Материалы Всесоюзн. конфер. Тбилиси. 1982. С.40.
Костюк П.Г. Современные проблемы физиологии и патологии нервной системы. // М.: Медицина. 1965. С.28-34
Кратин Ю.Г., Зубкова Н.А., Лавров В.В., Рассказовский С.П. Электрические реакции мозга кошки при дифференцировании световых стимулов. // Ж. высш. нервн. деят., 1975, т.25, №6. С.1203-1210.
Кратин Ю.Г. Электрические реакции мозга на тормозные сигналы. // Л.: Наука. 1967. 257с.
Кроль В.М., Специфика работы зрительных механизмов правого и левого полушарий мозга человека. // Ж. высш. нервн. деят, .1995, т.5, вып.6. С.1075-1084.
Кураев Г.А. Функциональная асимметрия коры мозга и обучение. // Ростов-на-Дону, Изд. Ростовского унив., 1982. 158с.
Кураев Г.А., Соболева И.В. Функциональная межполушарная асимметрия мозга и проблемы валеологии. // Валеология, 1996, №2. С.29-34.
Лавров В.В. Взаимодействие анализирующей и активирующей систем мозга кошки при анализе условных световых сигналов. // Автореф. дисс. канд. биол. наук // Ленинград, 1978. 23с.
Лебедев Б.А. К вопросу о парной работе больших полушарий мозга у человека. // Ж. высш. нервн. деят. 1954, т.4, №4. С.474-481.
Леушина Л.И., Невская А.А., Павловская М.Б. Асимметрия полушарий головного мозга с точки зрения опознания зрительных образов. // Сенсорные системы. Зрение. Л.: Наука. 1982. С.76-92.
Ливанов М.Н. О замыкании условных связей. // Электроэнцефалографическое исследование высшей нервной деятельности. // М.: Изд. АН СССР. 1962. С.174-186.
Ливанов М.Н. Пространственная организация процессов головного мозга. // М. Наука, 1972. 182с.
Лурия А.Р. Высшие корковые функции человека и их нарушения при локальных поражениях мозга. // М.: Изд. Московского унив., 1962. 432с.
Макаров Ф.Н. Комиссуральные связи двигательной коры у некоторых млекопитающих. // Ж. эволюц. биохим. и физиол. 1969, т.5, №5. С.507-512.
Макарова И.А. Характеристика асимметрии в двигательной коре мозга кошек. // Научные доклады высшей школы. Биол. н. 1979, 38. С.62-69.
Матова М.Л. Формирование асимметрии и симметрии зрительного восприятия в процессе практической деятельности человека. // Вопросы психологии. 1980, №1. С.64-72.
Меерсон Я.А. Проявление функциональной асимметрии полушарий головного мозга в осуществлении зрительно-гностических функций. // Физиология человека, 1996, т.22, №3. С.52-58.
Мнухина Р.С., Самойлова Л.А. К вопросу о механизме тормозящего влияния аминазина на замыкательную функцию коры. // Докл. АН СССР. 1970, т.191, №1. С.253-256.
Мосидзе В.М., Рижинашвили Р.С., Самадашвили У.В., Турашвили Р.Т. Функциональная асимметрия мозга. // Тбилиси: Мецниереба, 1977. – 120с.
Мосидзе В.М., Эзрохи В.Л. Взаимоотношения полушарий мозга. // Тбилиси: Мецниереба. 1986. 160с.
Наумова Т.С. Электрофизиологический анализ механизмов формирования условного рефлекса. // Л.: Медицина. 1968. 232с.
Овчинников Н.Ф. Симметрия – закономерность природы и принцип познания. // В кн.: Принцип симметрии. Историко-методологические проблемы. М.: Наука, 1978. С.5-46.
Павлова Л.П. Точилов К.С. К электроэнцефалографической характеристике парной работы больших полушарий человека при мышечной работе. // Физиол. ж. СССР. 1960, т.46, №7. С.777-785.
Печенкин А.А. Симметрия и структура познания. // Принцип симметрии. Историко-методологические проблемы. М.: Наука, 1978. С.89-121.
Подвигин Н.Ф., Куперман А.М., Миртов В.К., Чуева И.В. Описание изображений рецептивными полями сетчатки и наружного коленчатого тела кошки. // Проблемы нейрокибернетики. Ростов-на-Дону: РГУ. 1976. с.131.
Полюхов А.М. Моторная асимметрия мозга в позднем онтогенезе. // Ж. физиол. человека. 1982, т.8, №1. С.162-163.
Пятигорский Б.Я. Взаимодействие афферентных и нисходящих систем потоков на нейронах дорсального спинно-мозжечкового тракта. // Механизмы объединения нейронов в нервном центре. Л.: Наука. 1974. С.93-99.
Рабинович М.Я. Электрофизиологический анализ активности различных слоев коры больших полушарий при формировании условного рефлекса. // Ж. высш. нервн. деят., 1961. т.11, №3. С.463-468.
Рабинович М.Я. Нейрофизиология условной связи. // Механизмы формирования и торможения условных рефлексов. М.: Наука. 1973. С.95-114.
Сачков Ю.В. Развитие представлений физики об элементарных объектах в свете идей симметрии. // Вопросы философии, 1963, №2. С. 121-139.
Семенова Л.К. Развитие эндокринной системы у детей от рождения до 7 лет. // Биологический возраст и возрастная периодизация. М. 1978. С.96-101.
Сергеев Г.А., Павлова Л.П., Романенко А.Ф. Статистические методы исследования электроэнцефалограммы человека. //Л.: Наука. 1968. С.47-122.
Силаков В.Л. Регуляция центральных механизмов зрения. // Л.: Наука. 1982. 187с.
Симмерницкая Э.Г. Доминантность полушарий. // М.:Изд. МГУ. 1978. 95с.
Сметанкин Г.Н. Включение во временную связь нейронов зрительной и ассоциативной областей коры полушарий кошек. // Ж. высш. нервн. деят. 1970, т.20, №4. С.563-569.
Сологуб Е.Б. Электрическая активность мозга человека в процессе двигательной деятельности. // Л.: Медицина. 1973. 247с.
Трауготт Н.Н. Выявление эволюционных закономерностей в картине тотального полушарного и очагового выключения коры головного мозга человека. // 6 научное совещание и симпозиум по эволюционной физиологии, посвященные 90-летию со дня рождения Л.А. Орбели. Л.: 1972. С.211-212.
Трауготт Н.Н. О механизмах нарушения памяти. Доклад на чтениях, посвященных памяти академика Л.А. Орбели 30 октября 1968г. // Л.: Наука. 1973. 23с.
Труш В.Д., Ливанов М.Н. Оценка функциональной значимости биоэлектрических показателей с помощью управляющей ЭВМ. // Статистическая электрофизиология. Вильнюс. 1968. Ч.1. С.552-567.
Урманцев Ю.А. О природе правого и левого (основы теории диссфакторов). // Принцип симметрии. М.: Наука, 1978. С. 180-195.
Ухтомский А.А. Собрание сочинений, в 6-ти томах. 1954. Т.4.
Фарбер Д.А. Системная организация интегративной деятельности в онтогенезе ребенка. // Физиология человека. 1979, т.5, №3. С.516-521.
Фарбер Д.А., Дубровинская Н.В. Биологическая надежность и формирование функций ЦНС. // Научное совещание и симпозиум по эволюционной физиологии, посвященные 90-летию со дня рождения Л.А. Орбели. Л. 1972. С.217.
Фишман М.Н., Труш В.Д., Марковская И.Ф. Спектрально-корреляционный анализ ЭЭГ детей в норме и при задержке психического развития. // Физиология человека. 1983, т.9, №1. С.75-82.
Хананашвили М.М., Заркешев Э.Г., Силаков В.Л. Проявление эффекта обусловливания (выработки «временной связи») на нейронах коры, изолированной от подкорковых влияний. // Физиол. ж. СССР. 1971, т.57, №3. С.490-495.
Хрипкова А.Г., Фарбер Д.А. Основные направления исследований физиологии развития ребенка. Итоги и перспективы. // Физиология человека. 1983, т.9, №1. С.3-17.
Чораян О.Г. Информационная оценка вызванных импульсных потоков нервных клеток. // Биологическая и медицинская кибернетика. Ч.3. Нейрокибернетика. М. 1974. С.190-192.
Чуприков А.П. Особенности моторного доминирования у психически больных. // Нервно-психические заболевания экзогенно-органической природы. М. 1975. С.209-218.
Швырков В.Б. Нейрофизиологическое изучение системных механизмов поведения. // М.: Наука. 1978. 240с.
Швырков В.Б. Системная организация нейрофизиологических процессов в элементарном поведенческом акте. // Автореф. дисс. докт. мед. наук. М.: 1979. 38с.
Шевченко Д.Г. Активность нейронов зрительной коры в системных процессах смены поведенческих актов. // Нейрофизиологические механизмы поведения. М.: Наука. 1982. С.371-381.
Шульговский В.В., Котляр Б.И. Динамика активности нейронов зрительной коры в процессе формирования условно-рефлекторной оборонительной доминанты. // Ж. высш. нервн. деят., 1968, т.28, №5. С.799-806.
Щебланова Л.И. Межполушарная асимметрия ЭЭГ-реакций головного мозга человека при мыслительной деятельности. // Основные проблемы общ. возрастн. и пед. психол. М.: 1978. С.27-33.
Broca P. Remarques sur le siege de la faculte du language articule. // Bull. Soc. Anthrop., 1861, v.6. P.18-23. (Цитировано по Бианки В.Л. Индивидуальная и видовая межполушарная асимметрия у животных. // Ж. высш. нервн. деят., 1979, т. 29, № 2. С.295-303).
Bures J., Buresova O. (Буреш Я., Бурешова О. Межполушарная интеграция и механизмы памяти. // Механизмы формирования и торможения условных рефлексов. М.: Наука. 1973. С. 213-226.
Butler S.R., Glass A. Asymmetries in the electroencephalogramme associated with cerebral dominance. // Electroencephalogr. And Clin. Neurophysiol. 1974, v.36, №5. P.481-491.
Doty R., Nigrau N., Jamaga K. (Доти Р., Ниграу Н., Ямага К.) Односторонняя энграмма. // Основные проблемы электрофизиологии головного мозга. М.: Наука, 1974. С.171-178.
Gerstein G.L. Kiang N. An approach to the quantitative analysis of electrophysiological data from single neurons. // Biophys.J., 1960, №1. P.15-21.
Geschwind W. Specialization of the human brain. // Sci. Amer. 1979, v.241, №3. P.158-166.
Green P. Defective interhemispheric transfer in schizophrenia. // J. Abnorm. Psychol. 1978, v.87, №5. P.472-480.
Hecaen H. La symmetrie en neuropsychologie. // Totus homo. 1970, v.2, №1. P.8-15.
John E. (Джон Е. Статистическая теория обучения и памяти. // Механизмы формирования и торможения условных рефлексов. М.: Наука. 1973. С.183-213).
Johnson R.C., Cole R., Bowers J., Foiles S., Nikaido A., Patrick J., Woliver R. Hemispheric efficiency in middle and later adulthood. // Cortex. 1979, v.15, №1. P.109-119.
Jones B., Santi A. Lateral asymmetries in visual perception with and without eye movements. // Cortex. 1978, v.14, №2. P.164-168.
Katz A. Cognitive arithmetic: evidence for right hemispheric mediation in an elementary component stage. Quart. J. Exp. Psychol. 1980, v.32, №1. P.69-84.
Kornfeld C., Beatty J. EEG spectra during a long-term compensatory tracking task. // Bull. Psychonom. Soc. 1977. v.10, №1. P.46-48.
Olds J., Disterhoft J., Segal M., Hornblith C., Hirsh R. Learning centers of rat brain mapped by measuring latencies of conditioned unit responses. J. Neurophysiol. 1972, 35. P.202-212.
Osborne K., Gale A. Bilateral EEG differentiation of stimuli. // Bid. Psychol. 1976, v.4, №5. P.185-196.
Polich J.M. Left hemisphere supericrity for visual search. Cortex. 1980, v.16, №1. P.39-50.
Segundo J., Moor G., Stensaas L.,Bullock T. Sensitivity of neurons in Aplysiato temporal patterns of arriving impulses. // J. Exp. Biol., 1963, v.40. P.643-657.
Singer W. The effect of monocular deprivation on the cat parastriate cortex asymmetry between crossed and uncrossed pathways. // Brain Res. 1978, v.157, №2. P.351-355.
Taylor D.C. Differential rates of cerebral maturation between sexes and between hemispheres. // Lancet. 1969, v.2. P.140-142.
Webster W.G. Functional asymmetry between the cerebral hemispheres of the cat. // Neuropsychologia. 1972, v.10. P.75-87.