 Результаты
первого этапа исследования отражены на графиках (рис. 1)
Результаты
первого этапа исследования отражены на графиках (рис. 1)
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ МЕЖПОЛУШАРНОЙ АСИММЕТРИИ:
ЭКСПЕРИМЕНТАЛЬНЫЕ И КЛИНИЧЕСКИЕ АСПЕКТЫ ПРОБЛЕМЫ
Л.Л. Клименко
Институт химической физики РАН, Москва
Будучи актуальной мультидисциплинарной проблемой, возникшей в середине ХIХ века, функциональная межполушарная асимметрия (ФМА) и по сию пору содержит больше вопросов, чем ответов на них. Как феномен, ФМА достаточно хорошо изучена, отчасти понятна также ее биологическая роль: оптимизация процесса выбора при принятии решения путем ограничения числа степеней свободы; не оставляет сомнений, что полушария различаются по способу обработки информации, однако за границами феноменологии остается совокупность вопросов, касающихся материальных носителей этих различий – биохимических, элементоорганических, нейрофизиологических, энергетических и др. При этом центральным и полностью открытым к настоящему времени остается вопрос о цене – метаболической или энергетической – которую платит организм за формирование и поддержание ФМА. Меняется ли эта цена в процессе онтогенеза? Одинакова ли ФМА при развитии и старении? Исходя из представления о ФМА как о системе, важно понять закономерности отношений в иерархических уровнях системы и принципы обеспечения динамического взаимодействия между отдельными уровнями в норме, в процессе физиологического старения, а также при экстремальных воздействиях на организм, в том числе и при вовлеченности последнего в патологический процесс.
Таков краткий перечень задач, стоявших перед нами при исследовании ФМА как феномена и проблемы. Алгоритм исследования следующий.
Экспериментальная часть состоит из трех разделов.
В первом исследуется динамика показателей различных уровней ФМА: нейрофизиологического, биохимического, энергетического и поведенческого в процессе онтогенеза экспериментальных животных (при физиологическом старении).
Во втором рассматриваются межполушарные различия этих показателей у животных с разным типом ФМА (правшей, левшей и амбидекстров), отобранных по поведенческому и нейрофизиологическому критериям.
Третий раздел экспериментального исследования посвящен вопросу устойчивости структурно-функциональной организации межполушарной асимметрии при экстремальном воздействии, примером которого является радиационное облучение.
Клиническая часть исследования посвящена анализу роли функциональной межполушарной асимметрии в модуляции иммунного ответа организма при вовлеченности последнего в патологический процесс и изменению структурно-функциональной организации ФМА как при развитии патологического аутоиммунного заболевания, так и при экстремальном радиационном воздействии.
Экспериментальное исследование.
Нейрофизиологические, биохимические, поведенческие и энергетические паттерны при формировании и инволюции ФМА
Известно, что одним из валидных нейрофизиологических критериев ФМА является межполушарная разность уровня постоянного потенциала (УПП) головного мозга – это показатель типа и выраженности ФМА (В.Ф. Фокин с соавт, 1982; 1986). Было показано, что различие УПП в больших полушариях головного мозга крыс не менее чем на 2 мВ является признаком выраженной латерализации животного (правшества или левшества), а отсутствие этого межполушарного различия УПП свидетельствует об амбидекстрии (Л.Л. Клименко с соавт., 1987; 1998; 1999). Постоянные потенциалы милливольтного диапазона характеризуют стационарную систему управления (в отличие от оперативной, маркером которой является ЭЭГ, регистрирующая постсинаптические потенциалы микровольтного диапазона). Основной вклад в генерацию УПП вносят потенциалы гематоэнцефалического барьера и в меньшей мере мембранные потенциалы нервных и глиальных клеток; в связи с этим фактом УПП принято считать интегральным отражением мембранных потенциалов сосудистого и нейроглиального происхождения (R. Gumnit, 1974; G.G. Somjen et al. 1979; Дж. Дудел с соавт., 1985). Генерация мембранных потенциалов связана с энергозатратами, необходимыми для создания и поддержания ионных градиентов, функционирования К-Nа насоса и др. Данные экспериментальных исследований свидетельствуют о связи величины постоянного потенциала головного мозга с показателями энергетического обмена: с потреблением кислорода и глюкозы, накоплением гликогена, концентрацией АТФ и других энергетических метаболитов, а также с соотношением окисленных и восстановленных форм дыхательных ферментов NАD-NАDН и с интенсивностью реакций свободнорадикального окисления липидов мембран головного мозга (A.L. Hodgkin, 1964; S.D.Glick еt al., 1977; A. Lehmenkuhler, 1979; В.Ф. Фокин с соавт., 1994; Hansen A.J., 1985; F. Van Bel et al., 1998; Биохимия мозга под ред. И.П. Ашмарина, 1999). Совокупность этих данных позволяет рассматривать УПП как меру интенсивности энергетического метаболизма в ЦНС.
Согласно современным представлениям, рост, развитие и старение организма (приближение к конечному равновесному состоянию) сопровождается уменьшением энергетического метаболизма, определяемого по интенсивности потребления кислорода, концентрации АТФ в клетках или по энергетическому заряду Аткинсона (соотношение концентрации АТФ, АДФ и АМФ). При этом в мозге происходят характерные изменения его интегративной деятельности, одним из ведущих механизмов которой является функциональная межполушарная асимметрия (ФМА) (О.С. Адрианов, 1976; В.В. Фролькис, 1991; В.В. Абрамов, 1991).
Исследование нейрофизиологических, биохимических, энергетических и поведенческих характеристик в процессе формирования и старения системы ФМА было разделено на два этапа. Первый этап включал в себя определение моторной асимметрии, регистрацию и анализ УПП, измерение концентрации гидроперекисей, Шиффовых оснований фосфолипидов и липофусцина в полушариях головного мозга крыс разного возраста. На втором этапе исследования после определения моторной асимметрии у крыс регистрировали УПП в моторной области коры каждого полушария и параллельно измеряли температуру коры в той же области. На первом этапе работа была выполнена на 200 белых крысах - самцах в возрастном диапазоне от 3 до 34 месяцев, на втором этапе в эксперименте было использовано 53 крысы в возрастном диапазоне от 3 до 25 месяцев. Поведенческие эксперименты проводили в Т-образном лабиринте. Животных тестировали по побежкам: если из 10 побежек животное осуществляло 8 и более поворотов в одну сторону, его относили к группе, латерализованной по поведению - правшей или левшей, в зависимости от стороны поворота, остальных животных относили к амбидекстрам. Моторная асимметрия оценивалась по коэффициенту асимметрии, вычисляемому по формуле: Кас=|Ппр-5|, где Ппр- количество побежек направо.
Перед регистрацией УПП животных наркотизировали этаминалом натрия из расчета 0,34 мг на 100 г массы. После окончания острого опыта находящихся под наркозом крыс декапитировали.
Постоянные потенциалы отводили от твердой мозговой оболочки в лобных областях каждого полушария неполяризуемыми хлорсеребряными электродами с сопротивлением 10 кОм. Регистрировали УПП с помощью электрометра с входным сопротивлением 1014Ом. Характеристикой ФМА служила разность в значениях УПП в лобных областях каждого полушария Δ=(Fd-Fs), мВ.
Определение концентрации продуктов свободнорадикалъного окисления гидроперекисей (ГП) и Шиффовых оснований фосфолипидов (ШО) проводили в экстрактах фосфолипидов из каждого полушария в отдельности согласно методике, описанной ранее (Л.Л. Клименко с соавт., 1987). Концентрацию пигмента старения липофусцина определяли иначе - не в экстрактах фосфолипидов, а в гомогенатах коры головного мозга методом синхронного сканирования люминесценции (Я. Рабек, 1985) с помощью спектрофлуориметра Hitachi МРF-4. Параметром, характеризующим накопление липофусцина в коре, служило отношение интенсивности флуоресценции липофусцина (λmax=440 нм) к интенсивности флуоресценции триптофанила (λmax=340 нм). Таким образом, содержание липофусцина нормировалось на содержание белка в пробе. Температуру в моторной области коры полушарий измеряли с помощью констатан-манганиновых термопар, соединенных с компьютерно - измерительной системой "Аксамит" (Л.Л. Клименко, 2000).
Результаты
первого этапа исследования отражены на графиках (рис. 1)
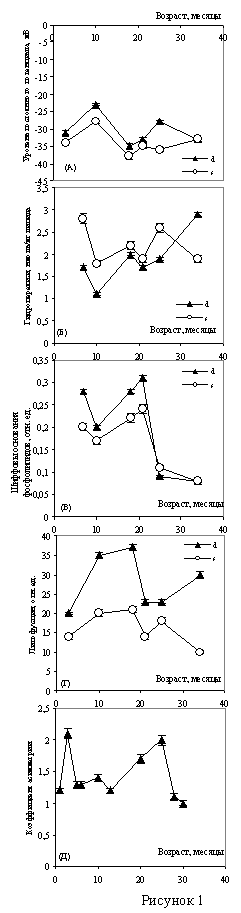
Рис. 1. Динамика уровня постоянного потенциала (а), гидроперекисей (б), Шиффовых оснований фосфолипидов (в) и липофусцина (г) в полушариях головного мозга крыс в процессе старения. Динамика коэффициента асимметрии поведения в Т-образном лабиринте (д).
Динамика уровня постоянного потенциала в возрастном диапазоне от 3 до 34 месяцев представлена М-образной кривой, имеющей два максимума. Постоянный потенциал головного мозга имеет максимальные значения в возрасте 10 и 28 месяцев, минимальные - в возрасте 3 и 21 месяц. Существенно при этом, что значения УПП в полушариях различаются в течение всего исследованного периода, т.е. в процессе онтогенеза между полушариями имеется существенная разница в интенсивности энергетического обмена (рис. 1 - а).
Динамика гидроперекисей, Шиффовых оснований фосфолипидов и липофусцина в полушариях головного мозга крыс представлена на графиках (рис. 1 - б, - в, - г). Все кривые имеют многофазный характер и концентрация исследованных продуктов ПОЛ различается в полушариях в течение всего периода онтогенеза. Графики, описывающие динамику ГП в полушариях, имеют два переломных момента, приходящиеся на молодой (10 мес.) и зрелый (20 мес.) возраст. Динамика конечных продуктов ПОЛ - Шиффовых оснований фосфолипидов также имеет сходный с динамикой ЛФ многофазный характер. Очевидное сходство динамических кривых ЛФ и ШО вполне объяснимо, так как в состав ЛФ входят соединения типа Шиффовых оснований, которые люминесцируют в той же области, что и ЛФ. Несмотря на традиционное представление о ЛФ, как о пигменте старения, накопление ЛФ - немонотонная функция возраста: скорость накопления различна в полушариях и максимальна в молодом возрасте. В правом полушарии скорость накопления ЛФ в период от 3 до 9 месяцев в 16,8 раза выше, чем в зрелом возрасте - от 9 до 20 месяцев. Такое большое различие в скорости накопления ЛФ в разные периоды онтогенеза ассоциируется как с высокой интенсивностью свободнорадикальных реакций - "в молодости старение идет быстрее" (Age pigments, 1981).,– так и с высокими затратами энергии, сопровождающими созревание мозга. Существует мнение, что ЛФ - это внутриклеточное депо кислорода (S. Bjorkerud, 1964; В.Н. Карнаухов, 1973), а его накопление и утилизация в процессе онтогенеза - это приспособление к гипоксии в молодом и старческом возрасте. Известно, что в процессе онтогенеза существует ряд критических периодов, связанных с созреванием мозга, когда значительно меняется его деятельность. Эти периоды соответствуют основным периодам жизни: достижения половой зрелости, репродуктивному и периоду угасания репродуктивной функции.
С этими периодами совпадают изменения в активности защитных ферментных систем, под контролем которых протекают процессы свободнорадикального окисления. Многофазность динамических кривых связана с соответствующими возрастными изменениями активности защитных ферментов. Ряд исследователей показал (K. Nandy, 1978; Tolmasoff J.М. et al., 1980; D. Roy et al., 1983), что СОД и ферментная система глутатион-трансфераза - глутатион-редуктаза в процессе онтогенеза претерпевают изменения активности, которые можно описать М-образной кривой, при этом переломные моменты этой кривой приходятся на те же возрастные периоды, что и в наших графиках. Объектом исследований возрастных изменений активности защитных ферментов был у этих авторов, как и в наших опытах, головной мозг крыс.
Многофазная динамика УПП, синхронизированная с таковой же всех исследованных продуктов, также находит объяснение в возрастных изменениях активности защитных ферментов. Действительно, изменение электрического сопротивления мембран и нарушение их барьерных функций, происходящие под влиянием свободнорадикального окисления липидов мембран головного мозга и под контролем защитных ферментов, не может не отразитъся на интегральном мембранном потенциале, каким является УПП.
Межполушарный различия, наблюдаемые во всех возрастных периодах и для всех продуктов свободнорадикального окисления, включая липофусцин, ассоциируется с представлением о различии в скорости старения полушарий. Благодаря тому, что в течение всего периода онтогенеза поддерживается концентрационный градиент свободных радикалов, можно сказать, что в полушариях головного мозга часы старения идут с разной скоростью.
Моторная асимметрия животных ранее, чем другие виды асимметрий привлекла внимание исследователей (R.L. Collins 1968; 1975; 1985; 1991; S.D. Glick et al., 1977; В.Ф. Фокин, 1982; J. Cenracec, 1991), однако механизм ее возникновения и связь с нейрофизиологическими, а тем более с биохимическими процессами, протекающими в полушариях головного мозга, долгое время не была исследована. Однако поведение – это высший иерархический уровень всего каскада адаптивных реакций, поэтому механизм возникновения моторной асимметрии, которая принимает самое непосредственное участие в осуществлении процесса выбора, нуждается в пристальном внимании.
На поведенческом уровне ФМА отражена в динамике коэффициента асимметрии, характеризующем поведение крыс в Т-образном лабиринте (рис. 1, д). Исходя из представления о сопряженности нейрофизиологических и биохимических процессов (постоянного потенциала и свободнорадикального окисления) в полушариях головного мозга, можно объяснить наличие двух максимумов (в 3 и 24 месяца) на графике зависимости коэффициента асимметрии от возраста. Эти возрастные периоды характеризуются недостаточной сбалансированностью процессов ПОЛ с процессами утилизации токсичных продуктов, связанной с низкой активностью ферментов защиты в молодом и позднем возрасте, что в свою очередь может приводить к большей биохимической и электрофизиологической асимметрии полушарий и, как следствие, к большей латерализации поведения. Корреляционный анализ выявил наличие достоверных связей между параметрами поведенческого, нейрофизиологического и биохимического уровней системы ФМА (Л.Л. Клименко 1986, Л.Л. Клименко с соавт, 1998).
Таким образом, поведенческий акт формируется под многоуровневым контролем, а взаимная сопряженность уровней, их иерархическое соподчинение и соблюдение количественных взаимоотношений между ними позволяет рассматривать ФМА как систему, обеспечивающую оптимизацию адаптивных реакций организма в течение всего периода онтогенеза.
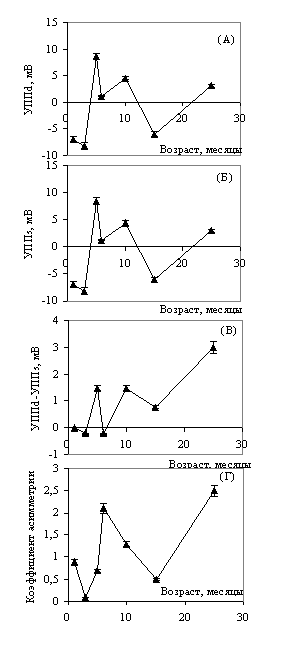
Рисунок 2.
Динамика уровня постоянного потенциала в правом (а) и левом (б) полушариях. Динамика асимметрии распределения постоянного потенциала (в) и коэффициента моторной асимметрии (г).
Очевидно, что поддержание межполушарной разности УПП, а также биохимических межполушарных различий не может происходить без соответствующих энергозатрат, таким образом, энергетический аспект формирования и поддержания ФМА оставался открытым. В связи с этим обстоятельством возник ряд вопросов. Что стоит за асимметрией распределения постоянного потенциала? Различается ли энергетический гомеостаз полушарий так же, как различаются параметры нейрофизиологического и биохимического уровней?
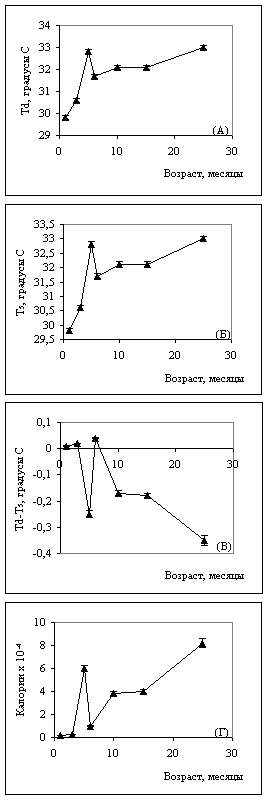
Рис. 3. Температурная динамика в правом (а) и левом (б) полушариях. Динамика температурной асимметрии (в) и энергозатрат мозга (д) в онтогенезе.
Таким образом, мы подошли к заявленному ранее вопросу о метаболической, а следовательно, об энергетической цене выбора. Для ответа на поставленные вопросы был выполнен второй экспериментальный этап исследования, при котором одновременно регистрировались УПП и температура в моторной области коры каждого полушария.
Результаты второго этапа представлены на графиках (рис. 2 и 3), из анализа которых следует: температура коры полушарий и УПП в процессе старения претерпевают синхронные многофазные изменения.
Динамику температуры, как и динамику УПП, можно разделить на три фазы: фазу быстрого роста (от 1 до 5 месяцев), фазу резкого уменьшения (от 5до 6 месяцев) и фазу стабилизации с постепенным ростом температуры и потенциала (от 6 до 25 месяцев) (рис. 2 (а, б); рис. 3 (а, б)). Эти периоды соответствуют определенным этапам формирования коры головного мозга: до 5 месяцев происходит образование новых клеток и связей, что сопровождается высоким уровнем энергообмена; в позднем возрасте вторичное повышение температуры связано с катаболическими процессами, идущими с высвобождением энергии.
Это повышение можно назвать пассивным, связанным с процессом утилизации энергии. Полученные динамические кривые в значительной степени совпадают с динамикой концентрации АТФ и потребления кислорода в различных органах и тканях белых крыс в процессе онтогенеза. При исследовании ректальной температуры мышей в процессе онтогенеза аналогичную температурную динамику авторы связывают с возрастным изменением гормонального статуса и метаболизма (M.J. Talan et al., 1986). Как видно из графиков, на которых представлены показатели температурной асимметрии полушарий и межполушарная разность потенциалов, УПП и температура в полушариях различаются во всех возрастных периодах (рис. 2- в; рис. 3- в). Этот факт можно рассматривать как показатель различной интенсивности энергообменных процессов в полушариях, поскольку оба исследуемых параметра являются мерой энергетического метаболизма.
Максимальных значений асимметрия распределения УПП, как и температурная асимметрия, достигает в 5 и 25 месяцев. В ряде работ (В.И. Пасечник, 1987; И.А. Шевелев с соавт, 1987) был проведен расчет теплопродукции (количества энергии), обеспечиваемой метаболизмом, в условной единице коры головного мозга (площадью 1мм2). Эти расчеты показали, что увеличение температуры коры в результате изменения метаболической активности или локального кровотока в элементарном объеме коры на 0,10 соответствует теплопродукции величиной 10-4 Вт или (10 х 0,239-4) калорий.
0,239х10-4 Кал х ΔТ°/ 0,1° (1)
Используя эти расчеты, можно определить энергетическую цену поддержания межполушарной асимметрии, изменяющуюся на протяжении всей жизни. Между постоянным потенциалом и температурой в различных возрастных группах существует достоверная корреляционная связь, максимальное количество связей приходится на возраст 15 месяцев. Понятно, что наибольшие энергетические траты приходятся на период наиболее выраженной межполушарной асимметрии.
Таким образом, очевидно, что информационные, энергетические и пластические процессы в ЦНС крыс связаны в единую систему межполушарной асимметрии. Параметры, характеризующие биологические уровни этой системы, синхронно и взаимосвязанно изменяются в процессе старения. Маркеры энергетического метаболизма коры больших полушарий головного мозга – постоянный потенциал и температура – претерпевают в позднем онтогенезе многофазные изменения, качественно сходные в обоих полушариях, но различающиеся количественно. Этот факт можно рассматривать как одно из объяснений асимметрии старения полушарий, показанной ранее. Выявленная многофазность динамики уровня постоянного потенциала, концентрации продуктов ПОЛ и температуры коры головного мозга ассоциируется со сменой типов регулирования обменных процессов и церебральных функций, обеспечивающих гомеостаз в разные периоды жизни.
Изложенные результаты свидетельствуют о том, что УПП является не только маркером интенсивности метаболизма и показателем энергетического гомеостаза мозга, но также характеризует тип межполушарной асимметрии, вносящей принципиальный вклад в организацию поведенческого акта. Таким образом, УПП представляет собой ключевой показатель, объединяющий нейрофизиологический, биохимический и поведенческий уровни межполушарной асимметрии.
При этом очевидно, что в формировании системы ФМА принимают участие и показатели других, более низких уровней биологической организации. Известно, что в энергетическом метаболизме мозга принимают участие металлоферменты, обладающие высокой биологической активностью.
Микроэлементы в нейрофизиологичских механизмах формирования межполушарной асимметрии
Высокая биологическая активность микроэлементов связана с их участием в синтезе витаминов и гормонов и их роли в качестве структурных единиц в молекулах ферментных систем (В.А. Дельва, 1965; Л.Р. Ноздрюхина, 1977; M.S. Black et al., 1981). Головной мозг, в частности кора больших полушарий, по содержанию микроэлементов занимает одно из первых мест среди других органов и тканей. Большинство микроэлементов (медь, алюминий, кремний, железо и др.), содержащихся в мозговой ткани, связаны с белками (K.S. Rajan, 1976), таким образом, микроэлементы участвуют во всем спектре метаболических процессов, что в полной мере относится и к метаболизму головного мозга (М. Диксон, Э. Уэбб, 1982). Белки, содержащие медь, преимущественно сосредоточены в коре головного мозга. Распределение микроэлементов в различных цитоархитектонических полях коры головного мозга человека неравномерно: выявлен более высокий уровень содержания микроэлементов в проекционных зонах различных анализаторов, что отражает значительную биохимическую активность этих высоко специализированных нервных образований (G.M. Gumming, 1948). При исследовании действия некоторых микроэлементов на функции высших отделов центральной нервной системы показано изменение основных процессов коркового возбуждения и торможения при введении в организм животных ионов кадмия, кобальта, железа, цинка и меди (В.С. Райцес, 1981). Существуют связанные с развитием концентрационные паттерны микроэлементов в коре и таламусе крыс, например у старых крыс более низкие концентрации алюминия, чем у молодых животных. Нarold Н. Sanstead (1986) провел широкое исследование влияния микроэлементов на деятельность мозга. R. Palm (1990) с соавторами выявил взаимосвязь между паттернами, определяющими хронологический возраст, и концентрацией Cu и Zn в различных отделах мозга.
N. Nakagawa (1998) обнаружил в гиппокампе мышей снижение концентрации Zn, Mn и Rb и повышение концентрации Cu при старении, что свидетельствует об участии именно этих микроэлементов в процессе старения мозга. В серии исследований, проведенных A. Takedа (1994–1999) с соавторами, выявлено, что изотопы Mn и Zn мозга проходят в кору мозга через гематоэнцефалический барьер, а роль ионов Mn в функции обонятельной системы и миндалины динамически связана с управляющими процессами в ЦНС. Этот же автор, исследуя изменение концентрации Mn в процессе старения крыс, показал, что в молодом возрасте требуется высокое содержание Mn и оно является критичным для нормального развития мозга.
Итак, рассмотренные нами микроэлементы оказывают принципиально важное влияние на развитие и функцию мозга. Однако все приведенные выше исследования были выполнены без анализа типа и выраженности асимметричной организации мозга. Исследование многоуровневой организации межполушарной асимметрии включало еще и анализ распределения микроэлементов в коре больших полушарий у животных с различным типом ФМА (правшей, левшей и амбидекстров).
У 30 крыс-самцов линии Вистар в возрасте 10 месяцев после тестирования в Т-образном лабиринте и измерения УПП, проводилась декапитация под наркозом. После экстирпации полушарий выделяли кору головного мозга. Затем с помощью эмиссионного спектрального анализа на спектрохимической системе GBC (Австралия) определяли концентрацию микроэлементов Cd, Co, Cu, Fe, Mn, Mo, P, S, и Zn в коре каждого полушария в отдельности.
Для анализа полученных результатов животные были разделены на 3 группы, различающиеся по моторной асимметрии: группу правшей, левшей и амбидекстров. При статистическом анализе было проведено сравнение концентрации микроэлементов в полушариях головного мозга у животных с разным типом ФМА и сравнение концентрации МЭ между полушариями внутри каждой из групп. Было показано, что в группе левшей достоверно выше концентрация кадмия, кобальта и меди в левом полушарии, чем в правом. Вероятно, при более интенсивном метаболизме субдоминантного полушария, чем доминантного, в нем значительно выше концентрация и активность специфических металлоферментов.
На рис. 4 графически представлено это различие, оцененное по критерию Стьюдента.
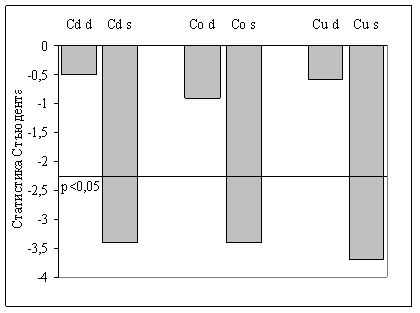
Рис. 4. Сравнение содержания микроэлементов (кадмия, кобальта и меди) в группах крыс левшей в правом (d) и левом (s) полушарии. На оси ординат отложены значения статистики Стьюдента. Пунктиром показан 5-процентный уровень значимости.
Межполушарное различие по кадмию и кобальту особенно важно, так как металлоферменты, активируемые этими ионами, принимают участие в формировании процессов возбуждения и торможения в ЦНС. Так, кадмий повышает критический уровень деполяризации мембран нервных клеток, а кобальт, входя в состав кобамидных ферментов, участвует в реакциях, протекающих с образованием свободных радикалов, что меняет электрогенные свойства мембран. Цианокобаламин (В 12) участвует в синтезе метильных групп и образовании холина.
Детальный анализ межполушарных различий в распределении микроэлементов был выполнен при помощи критерия Колмогорова-Смирнова, сравнивающего распределение МЭ в отдельных полушариях внутри групп и между ними.
В группе крыс правшей достоверно отличаются распределение молибдена и серы в полушариях. В правом полушарии распределение молибдена имеет крутой фронт и сосредоточено при более низких концентрациях, чем в левом - в нем распределение более пологое и концентрация молибдена выше. Можно предположить, что энергетический обмен, контролируемый флавопротеидными ферментами с молибденом в качестве активной группы, асимметричен в полушариях правшей.
В группе левшей показано достоверное межполушарное различие в распределении кадмия, марганца, серы. Вероятно, процессы окисления и восстановления, катализируемые марганцем, и процессы белкового синтеза протекают с разной интенсивностью в доминантном и субдоминантном полушариях левшей.
По критерию Колмогорова-Смирнова выявлено также различие в распределении микроэлементов в полушариях головного мозга у животных правшей и левшей. Достоверно отличаются эти группы по распределению кадмия, марганца, молибдена и серы – в левом полушарии (рис.5).
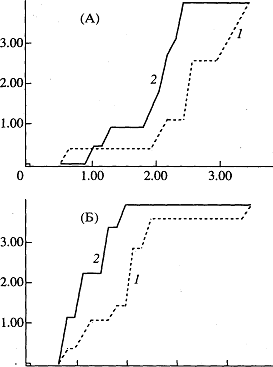
Рис. 5. Сравнение эмпирических функций распределения для содержания кадмия (А), марганца (Б) в левых полушариях правшей (1) и левшей (2). По оси абсцисс - концентрация, мкг/мл, по оси ординат - число случаев.
Таким образом, статистический анализ подтверждает положение о существовании единой системы межполушарной асимметрии, в которой низший (элементоорганический) и высший – нейрофизиологический и связанный с ним поведенческий уровень представляют собой единую иерархическую структуру.
Нейрофизиологические и биохимические межполушарные различия в группах с моторной и энергетической асимметриями
Получив представление о динамическом соотношении между уровнями системы функциональной межполушарной асимметрии, необходимо ответить на не менее актуальный вопрос о механизмах поддержания межполушарного нейрофизиологического и концентрационного градиента, обеспечивающего латерализацию поведения, в одном из возрастных периодов онтогенеза. Был выбран возрастной период 15 месяцев, где, как показано ранее, прослеживалась наибольшая плотность корреляционных связей между уровнями внутри системы ФМА.
Были сформированы четыре группы животных (в каждой из групп N=20) одинакового возраста – 15 месяцев, но отличающихся по поведению в Т-образном лабиринте (правши и левши по поведению) или по распределению УПП в полушариях головного мозга (левополушарные и правополушарные по УПП, с межполушарной разностью потенциала не менее 5 мВ). Результаты измерений приведены на гистограмме (рис.6).
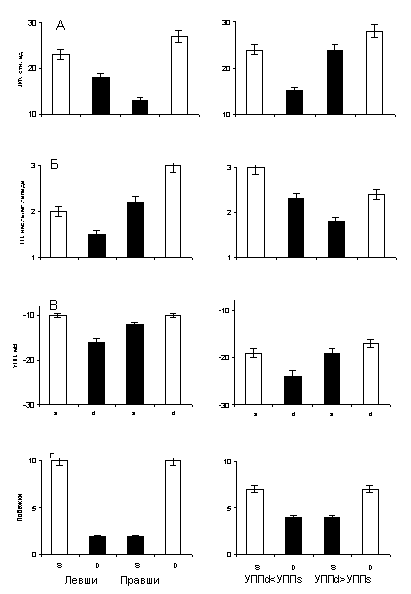
Рис.6. Гистограмма распределения показателей поведенческого, нейрофизиологического и биохимического уровней (побежки в Т-образном лабиринте; УПП, мВ; ГП, нМоль/мг и ЛФ, отн.ед., соответственно) в полушариях головного мозга крыс с выраженной асимметрией поведения и УПП. Доминантные полушария закрашены
На горизонтальных осях гистограммы условно представлены левши и правши. Индексами D и S (на оси «г») обозначены правая и левая части пространства. На оси ординат этой гистограммы отложено число побежек направо D в группах животных, латерализованых по поведению и потенциалу. На (а) - (в) гистограммы по оси абсцисс индексами d и s обозначены правое и левое полушария; по оси ординат – концентрация липофусцина (ЛФ) в отн. ед. и гидроперекисей (ГП) в нмоль/мг липида, а также значение УПП, мВ, соответственно в правом (d) и левом (s) полушариях.
В группах, сформированных по признаку асимметрии распределения потенциала (правая часть гистограммы), распределению потенциала соответствует определенная латерализация поведения: группа с превышением УПП справа обнаруживает тенденцию к «правшеству» (66% побежек направо), группа с превышением УПП слева обнаруживает тенденцию к «левшеству» (36% побежек направо). Это согласуется с существующим представлением о роли УПП в отражении функциональной моторной асимметрии. В группах правшей и левшей (левая часть рис. 6) значения УПП в доминантных полушариях меньше, чем в субдоминантных как минимум на 2±0,08 мВ. Известно, что высоким значениям УПП соответствует более высокий уровень метаболизма: утилизация глюкозы у человека выше в той области мозга, в которой выше УПП (Иванов К.П., 1972; Bures J., Buresova 0., Krivanek J, 1974; Hansen A.J, 1975; Илюхина В.А., Заболотских И.В., 1993; Шевелев И.А. и др., 1989). Таким образом, в субдоминантных полушариях, характеризующихся превышенным УПП по сравнению с доминантными, уровень метаболизма выше.
При рассмотрении биохимических параметров в отобранных группах животных становится очевидно, что концентрация ГП и ЛФ в полушариях также асимметрична: в доминантных полушариях их концентрация достоверно меньше, чем в субдоминантных, т.е. налицо концентрационный биохимический градиент в группах животных правшей и левшей, сформированных по различным критериям. Так, в правом, субдоминантном, полушарии у правшей концентрация ГП в два раза выше, чем в правом, доминантном, у левшей. Превышение концентрации ГП в субдоминантном полушарии по сравнению с доминантным в группах правшей и левшей составляет 31% и 21% соответственно. Концентрация ЛФ в субдоминантном и доминантном полушариях в группе правшей различается в 1,9 раза. Аналогичное соотношение между концентрацией продуктов ПОЛ в полушариях головного мозга наблюдается в группах, латерализованных по УПП. Так же как в группах правшей и левшей по поведению, в этих группах концентрация ГП в субдоминантном полушарии превышает их концентрацию в доминантном на 31%, а ЛФ - на 38%.
Итак, доминантные полушария в каждой из этих групп характеризуются меньшим УПП и меньшей концентрация продуктов ПОЛ по сравнению с субдоминантными – такова единая закономерность, прослеженная на всех уровнях.
Таким образом, латерализация поведения животных обеспечивается асимметрией процессов на нейрофизиологическом и биохимическом уровнях, т.е. наличием нейрофизиологических и биохимических различий.
Представление параметров всех уровней асимметрии в виде гистограммы продемонстрировало их четкое зеркальное отражение в правом и левом полушариях в группах с выраженной асимметрией поведения и постоянного потенциала. Такое зеркальное отражение показателей всех уровней в полушариях в группах, сформированных по различным критериям, не может быть случайным. Этот факт свидетельствует о взаимной сопряженности процессов различных уровней, их количественной взаимообусловленности и иерархической соподчиненности, т.е. об организации всех компонентов в единую функциональную систему межполушарной асимметрии.
Более высокая концентрация ЛФ в субдоминантном полушарии также ассоциируется с неодновременностью старения полушарий. Так как концентрация ЛФ – показатель не только хронологического, но и функционального возраста, то его меньшая концентрация в доминантном полушарии представляется биологически целесообразной. Действительно, почти двукратное превышение концентрации ЛФ в субдоминантном (правом полушарии у правшей и в левом у левшей) не может быть случайным. Поскольку хронологический возраст этих групп животных одинаков, концентрацию ЛФ в доминантном и субдоминантном полушариях можно рассматривать как показатель различного биологического возраста полушарий. Известный факт, что ЛФ является внутриклеточным депо кислорода, также хорошо соотносится с представлением о различной интенсивности метаболизма в субдоминантном и доминантном полушариях. Более высокая концентрация ЛФ в субдоминантном полушарии по сравнению с доминантным у крыс свидетельствует о более высоком уровне метаболизма, более высоком темпе старения именно субдоминантного полушария и соответственно о лучшей защищенности наиболее важного, доминантного полушария.
Статистически достоверные корреляционные связи объединяют все исследованные показатели: коэффициент асимметрии, УПП и концентрацию ГП и ЛФ. Структура корреляционных связей у правшей и левшей существенно отличается (Л.Л.Клименко, 1986).
Итак, взаимная сопряженность уровней, их иерархия и соблюдение количественных взаимоотношений между уровнями является условием, обеспечивающим доминантность полушария при организации целенаправленного поведения. Так как асимметрия поведения животных определяется, кроме всего прочего, и асимметрией биохимических процессов в полушариях головного мозга, истоки поведенческого акта можно проследить, начиная с регуляторных структурно-конформационных изменений мембраны, связанных с процессами свободнорадикального окисления и происходящих под контролем защитных ферментов.
Различия в температурных характеристиках правого и левого полушария у крыс
В ранее представленном выше разделе мы рассмотрели динамику температуры как показателя энергетического метаболизма коры больших полушарий головного мозга в позднем онтогенезе крыс. Однако открытым оставался вопрос о температурной асимметрии физиологически неравнозначных полушарий. Ранее выявленная рядом исследователей температурная неравнозначность полушарий не связывалась с их функциональной ролью и не рассматривалась как одно из проявлений функциональной межполушарной асимметрии (R. Busto et al., 1987; D.P. Cain et al., 1994). Из предыдущего раздела следует, что особенно велики межполушарные различия температуры в полушариях с различной функциональной ролью (доминантном и субдоминантном) у крыс с выраженной моторной преференцией – правшей и левшей (Л.Л. Клименко с соавт., 1998; 1999). Однако оставался неясным главный вопрос: существует ли различие между энергетическим метаболизмом мозга у крыс с разным типом ФМА - правшей, левшей и амбидекстров? Очевидно, что выявление температурной асимметрии коры полушарий, различающихся своей функциональной ролью, а также анализ связи между показателями энергозатратных процессов - постоянным потенциалом и температурой - может пролить свет на энергетический аспект формирования ФМА.
Для анализа полученных результатов все животные были разделены на группы, отличающиеся по независимым критериям: по поведенческому критерию – моторной асимметрии и по нейрофизиологическому – асимметрии распределения постоянного потенциала в полушариях - так называемые правши, левши и амбидекстры по поведению и по УПП. К правшам по поведению относили животных, которые в Т-образном лабиринте делали от 7 до 10 поворотов направо (N=11), к левшам – делавших от 0 до 3 поворотов направо (N=13) к амбидекстрам - тех, кто делал от 4 до б поворотов направо (N=29). Разделение животных на группы по поведению проводилось в широком диапазоне (отбирались животные не только с ярко выраженной латерализацией поведения, но и проявляющие тенденцию к латерализации) для того, чтобы исследовать возможные проявления температурной асимметрии у слабо латерализованных животных. При разделении на группы по распределению потенциала к правшам относили животных, у которых значение потенциала в левом, доминантном, полушарии было меньше, чем в правом, субдоминантном, на 2 мВ (N=22), к левшам - тех, у которых постоянный потенциал был ниже на 2 мВ в правом полушарии (N=14), к амбидекстрам относили животных, у которых межполушарная разность потенциала не превышала 1 мВ (N=17). Подобная идентификация типов асимметрии по нейрофизиологическому критерию совпадает с идентификацией по поведенческому критерию: правши, левши и амбидекстры, разделенные на группы по распределению постоянного потенциала, демонстрируют в Т-образном лабиринте поведение, соответствующее данному типу асимметрии. Достоверных различий по возрасту во всех указанных группах выявлено не было: возрастной состав групп был одинаков и составлял в среднем 5±0,2 месяцев. (Рис.7).
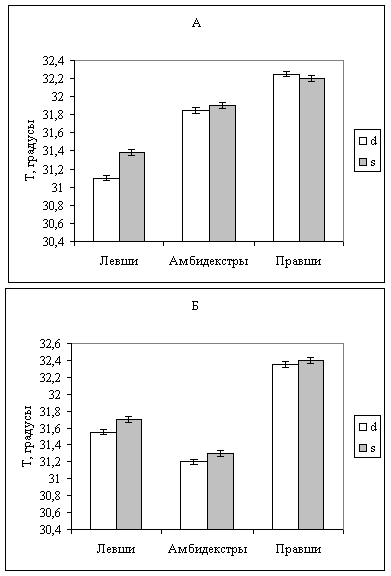
Рис. 7. Температура моторной области коры больших полушарий головного мозга крыс – правшей, левшей и амбидекстров. Группы сформированы по поведенческому (А) и по нейрофизиологическому (Б) критериям. d-правое полушарие; s-левое полушарие
Сравнение средних значений температуры моторной области коры полушарий проводилось с помощью непараметрических критериев и в несколько этапов. Средние значения температуры (по критерию Вилкоксона - Манна - Уитни) в группах крыс – правшей, левшей и амбидекстров, отобранных по поведению и потенциалу представлены на рис.7- а и -б. В группах, сформированных по поведенческому критерию, выявлено близкое к достоверному различие между температурой мозга правшей и левшей: правое полушарие правшей на 1,19±0,02 градуса, а левое – на 0,88±0,01 теплее, чем одноименные полушария левшей (рис. 7-а). В группах, сформированных по нейрофизиологическому критерию, достоверно различается температура мозга правшей и амбидекстров: правое полушарие правшей на 1,24±0,03 градуса, а левое – на 1,21±0,02 градуса теплее, чем у амбидекстров (рис. 7-6), что ассоциируется с ранее высказанным представлением об амбидекстрии как наименее функционально активном состоянии ЦНС, характеризующимся низким уровнем энергообмена.
При сравнении с помощью критерия Вальда-Вольфовица различия температуры между полушариями в группах крыс – правшей, левшей и амбидекстров, сформированных как по поведенческому, так и по нейрофизиологическому критерию, было показано, что температура между полушариями в группе левшей и в группе амбидекстров достоверное различается (р<0,05) (рис.8). Левое полушарие у левшей оказалось теплее на 0,3±0,01 градуса, чем правое, а у амбидекстров - на 0,08±0,02 градуса теплее, чем правое.
В группах правшей и амбидекстров по поведению с помощью того же критерия было выявлено достоверное (р<0,05) различие температуры между полушариями; при этом температура в левом полушарии правшей была выше на 0,08+0,01 градуса, а в левом полушарии амбидекстров – выше на 0,11±0,02 градуса, чем в правом. Очевидно, более высокая температура левого полушария соответствует более высокому уровню энергообменных процессов в нем.
Сравнение средних значений температурной асимметрии полушарий в разных группах с помощью критерия Вилкоксона-Манна-Уитни показало наличие достоверных различий этого параметра у правшей и левшей по поведению: температурная асимметрия левшей на порядок больше, чем у правшей (рис.8-а).
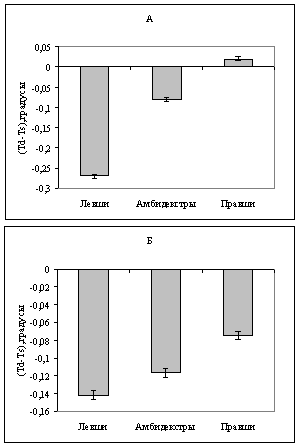
Рис.8. Температурная межполушарная асимметрия крыс-правшей, левшей и амбидекстров. Группы сформированы по поведенческому (А) и по нейрофизиологическому (Б) критериям.
Таким образом, температурная асимметрия полушарий в группах, сформированных по поведению, максимальна у левшей, минимальна - у правшей, амбидекстры занимают промежуточное положение. В группах, сформированных по потенциалу, максимальная и близкая по значению температурная асимметрия выявлена в группах левшей и амбидекстров, а минимальная - также у правшей (рис. 8-б). Температурная асимметрия полушарий правшей, левшей и амбидекстров, как видно из рисунка 8, практически не зависит от критерия, по которому формируются группы животных (по поведению или по потенциалу) и имеет близкие значения в одноименных группах.
Таким образом, выявлен не только температурный градиент между полушариями, но и его различие у животных с разным типом ФМА. Высокие значения температурной асимметрии у левшей и амбидекстров, по нашему мнению, могут свидетельствовать о более высоких энергетических затратах организма на поддержание функциональной межполушарной асимметрии у животных именно с этими видами асимметрии.
Таким образом, в результате проведенного исследования выявлен межполушарный температурный градиент, различающийся у крыс с разным типом межполушарной асимметрии. Оба показателя энергетического метаболизма – постоянный потенциал и температура коры больших полушарий – связаны между собой достоверными корреляционными связями, структура которых у правшей, левшей и амбидекстров различна (Л.Л.Клименко с соавт., 1999).
Температурная асимметрия полушарий, достоверно различающаяся у животных с разным типом ФМА, свидетельствует о том, что в основе ранее выявленных межполушарных асимметрий – биохимической, нейрофизиологической, элементоорганической - лежит различие в интенсивности энергетического метаболизма полушарий ("асимметрия метаболизма").
Итак, итогом экспериментального раздела исследования являются следующие основные положения: ФМА – это многоуровневая иерархическая система, внутри которой, по выражению фон Берталанфи, в свою очередь существует «система переплетающихся иерархий», т.е. нейрофизиологические, биохимические, энергетические и поведенческие корреляты ФМА находятся между собой в динамическом взаимодействии, их возрастная динамика синхронизирована и взаимообусловлена, а структура связей внутри системы меняется в пространстве и времени. Однако остается открытым вопрос об устойчивости этой системы. До сих пор мы рассматривали ее изменение в процессе нормального физиологического старения, в следующем разделе мы рассмотрим особенности структурно-функциональной организации асимметрии при ускоренном (радиационном) старении.
Структурно-функциональная организация функциональной асимметрии при ускоренном (радиационном) старении
Итак, феноменология старения и инволюции функциональной межполушарной асимметрии (ФМА) включает в себя возрастные изменения показателей различных уровней: поведенческого, нейрофизиологического, биохимического, молекулярного, энергетического и других. Изучение взаимосвязи между этими уровнями в процессе позднего онтогенеза дает представление о старении системы ФМА в целом.
Исследуя системную организацию ФМА и ее изменение в процессе старения, мы задались вопросом: насколько прочны количественные и иерархические отношения между различными уровнями системы, сохраняется ли структурно-функциональная организация ФМА при неблагоприятных воздействиях на организм, например, когда нормальное, физиологическое старение, становится патологическим, ускоренным?
Известно, что кроме естественного (физиологического) старения с различными целями исследуется "ускоренное" старение, которое вызывается неблагоприятными воздействиями на организм. Чаще всего это облучение, или температурный шок, или разного происхождения стрессы (Wheller K.T. et al, 1981). Преждевременное старение может быть "ускоренным" по отношению к естественному, а также обладать специфическими чертами. Общепринятым является мнение, что причинные события, определяющие скорость старения, происходят в постмитотических клетках, в особенности в клетках центральной нервной системы ввиду ее важной интегрирующей роли в организме. В качестве модели патологического старения мы использовали ускоренное "преждевременное" старение, вызываемое радиационным воздействием на организм. При использовании этой модели программа старения реализуется быстрее на 15-20%, чем при нормальном старении, однако структура вариационного ряда продолжительности жизни не меняется: т.е. кривые выживания остаются стандартными, они совмещаются в относительных координатах. При ускоренном старении не наблюдается каких-либо патологических соматических повреждений, но в клетках печени и головного мозга возникают дефекты структуры ДНК, опознаваемые нуклеазой S1, характерные для нормального старения ( Потапенко А.И. с соавт., 1988; А.И. Потапенко, Л.К. Обухова, 1992).
Исследование структурно-функциональной организации ФМА при ускоренном радиационном старении проходило в несколько этапов.
В ранее проведенном исследовании было показано изменение моторной асимметрии мышей-самок (N=40) линии СВА в возрасте от 5 до 29 месяцев, подвергнутых рентгеновскому облучению дозой 10 Гр. Использовалась рентгеновская установка РУТ-200-20-3 мощностью 2.5 Гр/мин. Облучали только голову животного, тело было закрыто стальным экраном толщиной 9 мм. Перед облучением вводили подкожно нембутал в дозе 40 мг/кг массы тела. В зависимости от индивидуальной чувствительности состояние наркоза длилось от 0.5 до 3 ч. В результате исследования было показано изменение моторной асимметрии в контрольной и облученной группе, причем кривые динамики асимметрии в обоих группах являются зеркальным отображением: в то время как у интактной группы в возрасте 12-15 мес. наблюдается максимальное значение Кас, у облученных животных на тот же возрастной период приходится минимум. В облученной группе наблюдается увеличение количества крыс-левшей в 1,5 раза, т.е. облучение приводит к отчетливо выраженному полевению популяции. Значения Кас в точках экстремумов близки и составляют 3,4 для интактной когорты и 3,1 - для облученной. Динамика моторной асимметрии мышей аналогична таковой же у крыс, отличие состоит лишь в том, что экстремальное значение Кас у крыс приходится на более поздний возраст 25 мес, а у мышей этот экстремум достигается в возрасте 14-16 мес. Периоды эквипотенциальности мозга в возрасте 5-10 мес. соответствуют снижению половой функции мышей и глубокой старости (больше 20 мес.). Четко выраженная латерализация в средних возрастах не связана с вымиранием популяции: в это время все животные были живы (Л.К. Обухова, Л.Л. Клименко, А.С. Соловьева, 1997). Парадокс в том, что максимум не связан с физиологическим расцветом (2-5 мес.), он приходится на возраст близкий к средней продолжительности жизни. Известно, что наибольшая продуктивность творческой деятельности людей также отмечена в среднем возрасте. Ее динамика удивительным образом напоминает кривую динамики моторной асимметрии, приведенную на рис. 1-д: значительный максимум в 40-45 лет, спад и второй меньший максимум после 60 лет (Д. Пельц, Ф. Эндрюс, 1973).
Наиболее естественной причиной инверсии поведения следует считать радиационное повреждение мозга после облучения дозой 10 Гр. При этом более чувствительным к радиационному повреждению оказалось доминантное левое полушарие.
Сравнение динамики УПП и липофусцина в полушариях головного мозга при нормальном и ускоренном старении
Исследовав динамику межполушарной моторной асимметрии при ускоренном старении, мы показали, что у ускоренно стареющих мышей моторная асимметрия отличается от контрольной увеличенным количеством левшей, т.е. при ускорении старения происходит инверсия типа межполушарной асимметрии. Такое "полевение" популяции можно рассматривать как показатель неодинаковой скорости старения полушарий, наиболее отчетливо проявляющейся при ускорении старения. Однако более глубокие причины инверсии ФМА при ускоренном (радиационном) старении нуждались в дополнительном исследовании. Чтобы внести ясность в понимание причин инверсии ФМА на следующем этапе было проведено сравнение показателей нейрофизиологического и биохимического уровней – УПП и липофусцина при нормальном и ускоренном старении.
В работе было использовано 70 мышей-самок линии СВА в возрасте 2, 7, 14 и 26 месяцев. Всего было 5 групп мышей по 10 животных в каждой группе. В возрасте 2 месяца была одна контрольная группа, в возрасте 7 месяцев было две группы контрольных мышей и одна группа – экспериментальных. В возрасте 14 месяцев была одна группа контрольная, другая – с экспериментальными (облученными) животными и в возрасте 26 месяцев была одна группа с контрольными животными.
После измерения УПП в полушариях головного мозга определяли концентрацию липофусцина в гомогенатах коры с помощью метода синхронного сканирования люминесценции. На представленном графике (рис. 9) видно, что в процессе нормального старения мышей УПП претерпевает многофазные изменения, близкие по типу к динамике потенциала при нормальном старении крыс (рис. 9, верхний график).
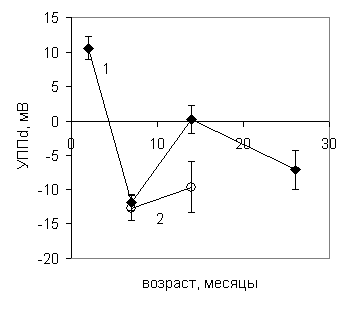
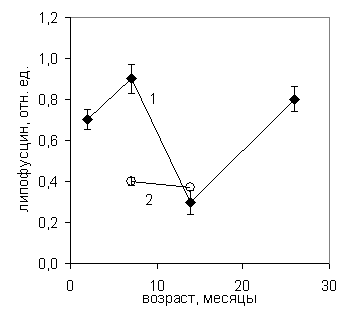
Рис.9 .Динамика уровня постоянного потенциала (вверху) и концентрации липофусцина (внизу) в правом полушарии головного мозга мышей при нормальном (1) и ускоренном (2) старении.
В раннем возрасте (2 мес.) значение УПП максимально, в возрасте 7 и 26 месяцев УПП имеет минимальные значения. Показатели УПП в экспериментальных группах достоверно отличаются от контрольных. Снижение УПП наблюдается как в возрасте 7 месяцев, так и в возрасте 14 месяцев.
Особенно сильно это снижение выражено в возрасте 14 месяцев: у экспериментальных животных УПП снижен в 37,4 раза, что ассоциируется как с представлением об энергодефиците позднего возраста, так и с пострадиационным энергодефицитом. Межполушарная разность потенциала сохраняется в течение всего периода онтогенеза (рис.10, верхний график), однако у ускоренно стареющих животных она радикально отличается от контрольных.
1 1 2 2
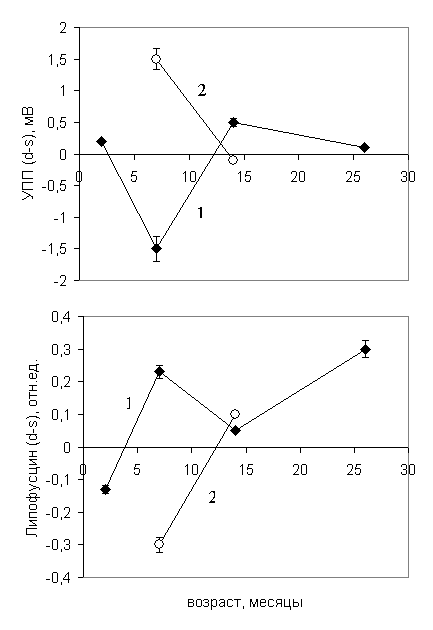
Рис.10. Динамика межполушарной разности потенциала (вверху) и межполушарной разности липофусцина (внизу) при нормальном (1) и ускоренном (2) старении.
Особенно сильно это отличие выражено в возрасте 7 месяцев: при ускорении старения межполушарная разность потенциалов УПП (d-s) мВ на 106% больше, чем в контроле! Таким образом, мы наблюдаем инверсию межполушарных различий, иначе говоря, если в контроле у большинства животных в возрасте 7 месяцев доминантным было левое полушарие, определяющее правостороннюю поворотную тенденцию, то при ускорении старения доминантным стало правое полушарие, что привело к показанному ранее "полевению" популяции.
Динамика концентрации липофусцина в полушариях головного мозга мышей при нормальном старении также синхронизирована с изменением УПП и носит многофазный М-образный характер (рис. 10, нижний график). Концентрация ЛФ максимальна в 7 и 26 месяцев и минимальна в 14 месяцев. Межполушарная разность концентрации липофусцина – биохимический градиент – также сохраняется в течение всего периода онтогенеза, при этом концентрация ЛФ в правом, субдоминантном для большинства животных, полушарии остается выше, чем в левом, почти во всех возрастных группах, кроме группы двухмесячных мышей (рис. 10, нижний график). Этот результат подтверждает сделанные ранее выводы о том, что накопление ЛФ – немонотонная функция возраста и различная концентрация его в полушариях свидетельствует о различии в скорости старения полушарий. Поскольку ЛФ является внутриклеточным депо кислорода, его накопление и утилизация в процессе онтогенеза может рассматриваться как приспособление к гипоксии в молодом и старческом возрасте. Как известно, ЛФ представляет собой одну из адаптивных метаболических систем, поэтому динамика его концентрации в онтогенезе отражает возрастные изменения функционального состояния и энергозатрат головного мозга и сбалансирована с активностью ферментов антиоксидантной защиты. Аналогично полученным ранее результатам исследования динамики потенциала при нормальном старении крыс, УПП уменьшается при снижении сопротивления мембран нервных и глиальных клеток, (увеличении проницаемости мембран) сопровождающим процессы ПОЛ. Сходство динамики УПП с динамикой активности защитных ферментов также свидетельствует о том, что все исследованные процессы являются составными частями единой адаптивной системы.
Концентрация ЛФ в полушариях головного мозга ускоренно стареющих, под влиянием радиационного облучения, мышей снижена в два раза в возрастной группе 7 месяцев. В то же время у экспериментальных животных в 14 месяцев концентрация ЛФ не отличается от контроля. Таким образом, на примере ЛФ можно наблюдать возрастные различия скорости старения полушарий головного мозга. Можно предположить, что уменьшение концентрации ЛФ у экспериментальных животных в возрасте 7 месяцев связано с высокой антиоксидантной активностью ферментов, характерной для этого возраста.
Как видно из рисунка, межполушарная разность концентрации ЛФ (d-s) в возрасте 7 месяцев у экспериментальных животных снижается, переходя в отрицательное значение, таким образом, наблюдается инверсия биохимического концентрационного межполушарного градиента, так же, как и нейрофизиологического, т.е. межполушарной разности потенциалов. Очевидно, что биохимическая инверсия является одной из причин поведенческой инверсии, т.е. лежит в основе "полевения" популяции животных после применения модели ускоренного старения.
С помощью корреляционного анализа в контрольных группах разного возраста были выявлены достоверные корреляционные связи между УПП и ЛФ, подтверждающие положение о единой многоуровневой системе межполушарной асимметрии (таблица 1). Однако в группах ускоренно стареющих животных корреляционные связи между показателями нейрофизиологического и биохимического уровней отсутствуют: ускорение старения приводит к разрыву связей, т. е. нарушению согласованности между уровнями ФМА, что ассоциируется с возрастным нарушением интегративной деятельности мозга.
Таблица 1.
Корреляционные связи между постоянным потенциалом и концентрацией липофусцина в коре больших полушарий головного мозга здоровых мышей 14 месяцев
|
|
ЛФs |
ЛФ(d-s) |
|
УППd |
-0,76 р=0,01 |
0,70 р=0,021 |
|
УППs |
Нет корреляции |
0,64 р==0,044 |
|
УПП(d-s) |
-0,78 р==0,008 |
Нет корреляции |
Таким образом, проведенное исследование показало, что многофазная возрастная динамика уровня постоянного потенциала синхронизирована с многофазной динамикой липофусцина, а ускорение старения приводит к снижению постоянного потенциала, что ассоциируется с возникновением радиационного энергодефицита в ЦНС; ускорение старения, под влиянием облучения, вызывает также уменьшение концентрации липофусцина в полушариях головного мозга; межполушарная разность потенциалов, как и межполушарная концентрационная разность липофусцина при ускорении старения претерпевает инверсию, что приводит впоследствии к поведенческой инверсии – "полевению" популяции экспериментальных животных.
Сравнение динамики уровня постоянного потенциала и микроэлементов в полушариях головного мозга мышей при нормальном и ускоренном старении
Как было показано ранее, концентрация микроэлементов, вносящих существенный вклад в формирование системы межполушарной асимметрии, достоверно различается в физиологически неравнозначных (доминантном и субдоминантном) полушариях головного мозга крыс. Кроме того, концентрация МЭ связана статистически значимыми корреляционными связями с показателем нейрофизиологического уровня – УПП, который одновременно является и маркером энергетического метаболизма мозга. Таким образом, концентрация микроэлементов является показателем элементоорганического уровня, являющегося дополнительным структурным элементом в иерархии системы ФМА. Известно участие МЭ в функциональной активности мозга, также как и в механизме старения мозга. Однако участие микроэлементов в феномене старения системы функциональной межполушарной асимметрии нуждается в дальнейшем исследовании. Анализ состояния системы ФМА при ускоренном старении дает представление об устойчивости системы к такому неблагоприятному воздействию, как радиационное облучение.
В экспериментальном исследовании было использовано 68 мышей-самок линии СВА в возрасте 14 и 26 месяцев. 32 контрольных и 36 экспериментальных животных были разделены на 4 группы. Контрольные мыши: возраст 14 месяцев (N=17) и 26 месяцев (N=15); экспериментальные мыши: возраст 14 месяцев (N=17) и 26 месяцев (N=19).
Моторная асимметрия исследовалась по поведению в Т-образном лабиринте, измерение УПП проводили традиционным методом. После окончания острого опыта находящихся под наркозом мышей декапитировали.
В экспериментальных группах животных определение межполушарной асимметрии мышей по двум критериям – поведенческому (побежкам в Т-образном лабиринте) и нейрофизиологическому (межполушарному градиенту УПП) – было проведено дважды: до и после облучения.
Для исследования концентрации микроэлементов в коре головного мозга применялся эмиссионный спектральный анализ. Определяли концентрацию Al, Cd, Co, Cu, Fe, Mg, Mn, Mo, P, Pb, S, Zn. Нижний предел концентрации в зависимости от элемента составляет от 1 х 10 до 1 х 103 мкг/мл. Инструментальная точность определения была не ниже 5% от определяемой концентрации. Количественно микроэлементы оценивались в мкг/100 мг мозговой ткани.
Анализ средних значений исследованных параметров в контрольных группах показал снижение концентрации меди в полушариях головного мозга в возрасте 26 месяцев по сравнению с 14-ю месяцами: в 14 месяцев в правом полушарии концентрация меди составляла (6,193±0,035) мкг/100 мг, а в 26 месяцев концентрация меди снизилась до значения (5,123±0,028) мкг/100 мг. При сравнении контрольных и экспериментальных групп наблюдаются многочисленные достоверные различия.
Статистика сравнения проводилась по критериям Стьюдента и Вилкоксона-Манна-Уитни; по всем критериям получены сходные результаты.
При сравнении средних значений в контрольных и экспериментальных группах было показано, что средние значения моторной асимметрии мышей в возрасте 26 месяцев до и после облучения достоверно различались: у облученных мышей количество побежек направо снижается, т.е. популяция экспериментальных мышей "левеет", что свидетельствует о различной чувствительности к облучению полушарий головного мозга (таблица 2).
Таблица 2
Изменение моторной и нейрофизиологической асимметрии мышей в возрасте 26 месяцев до и после облучения
|
|
Контроль, возраст 26 месяцев. |
Облучение, возраст 26 месяцев |
|
Побежки в Т-образном лабиринте |
5±0,4 |
2,8±0,3 |
|
УППd, мВ |
7,1±1,0 |
-8,07±1,05 |
|
УППs, мВ |
7,7±1,1 |
-5,5±1,03 |
|
УПП(d-s) ,мВ |
-0,26±0,6 |
-2,5±0,4 |
Подобное полевение популяции облученных мышей наблюдали мы и в предыдущем эксперименте. Снижение средних значений УПП наблюдается при облучении как в возрасте 14, так и 26 месяцев, что можно объяснить радиационным усилением свободнорадикального окисления и связанным с ним увеличением проницаемости мембран и падением мембранных потенциалов, интегральной характеристикой которых является УПП. Аналогичное снижение УПП в позднем онтогенезе наблюдали мы ранее при физиологическом старении крыс. Как видно из таблицы 2, при облучении наблюдается увеличение отрицательного значения межполушарной разности потенциалов, которая, как известно, является нейрофизиологической характеристикой ФМА: таким образом, УПП в правом полушарии снижается гораздо больше, чем в левом, по нейрофизиологическому критерию это полушарие стало доминантным, что повлекло за собой выявленную инверсию моторной асимметрии – полевение популяции.
Средние значения концентрации микроэлементов. Сравнение всей контрольной группы со всей экспериментальной, без деления на подгруппы, отличающиеся по возрасту и латерализации, выявило достоверные концентрационные различия как в правом, так и в левом полушарии: концентрация Al, Cu, Mg, Mn и Zn в контроле выше, чем у облученных животных.
В различных возрастных контрольных и экспериментальных группах также выявлены достоверные концентрационные различия. В 14 месяцев у ускоренно стареющих мышей в обоих полушариях наблюдается снижение концентрации Al и Zn. В 26 месяцев в левом полушарии у облученных мышей снижаются концентрации Al и Mn, а в правом полушарии кроме снижения концентрации этих же микроэлементов, выявлено также снижение концентрации Cu и Mg (рис. 11).



Рис. 11. Изменение концентрации микроэлементов в правом полушарии головного мозга мышей при ускоренном старении (контроль – серого цвета, ускоренное
Подобное возрастное снижение микроэлементов в различных отделах мозга наблюдали ранее N. Nakagava (1998) и A. Takeda (1999); таким образом, в связи с жизненно важной биологической ролью микроэлементов, их участие в возрастном изменении метаболизма является ключом к пониманию механизма старения мозга.
Сравнение концентрации микроэлементов в правом и левом полушариях у контрольных и экспериментальных мышей было проведено после предварительного разделения групп в возрасте 26 месяцев на правшей и левшей по моторной асимметрии. Сравнение концентраций микроэлементов проводили между идентичными полушариями правшей и левшей. В контрольных группах не было выявлено достоверных концентрационных различий между сравниваемыми полушариями, однако в группе облученных существует достоверное различие в концентрации Zn – в правом (доминантном) полушарии левшей концентрация Zn меньше, чем в правом (субдоминантном) полушарии правшей: (18,7±0,07) мкг/100 мг у левшей и (23,6±0,03) мкг/100 мг у правшей; таким образом, можно говорить о различном слдержании МЭ в функционально неравнозначных полушариях.
Между показателями различных уровней существуют многочисленные достоверные корреляционные связи, что ассоциируется с объединением рассматриваемого нами в данном исследовании элементоорганического уровня в единую систему функциональной межполушарной асимметрии с характерными для понятия "система" причинно-следственными и иеррархическими отношениями между уровнями. Однако общее количество корреляционных связей, которое можно рассматривать в качестве оценки степени согласованности изменений сравниваемых величин, различается в полушариях головного мозга у контрольных и экспериментальных животных.
После облучения происходит перераспределение связей, заключающееся в частичном распаде связей между всеми исследованными показателями и выравнивании количества связей в правом и левом полушариях. Так, в левом полушарии в контроле выявлены достоверные связи между Zn и Fe; Zn и Al, а при облучении эти связи отсутствуют, но возникает связь между Zn и Cu, Zn и Mg. Корреляционная связь между Cu и Mg остается неизменной в контроле и в экспериментальной группе, а связь между Mn и Cu возникает лишь при облучении. Аналогичная картина наблюдается и в правом полушарии. Уменьшение числа (или распад) достоверных корреляционных связей между микроэлементами при ускоренном старении соответствует ранее полученным данным о распаде корреляционных связей при нормальном старении системы межполушарной асимметрии (Л.Л. Клименко с соавт., 1994). Выравнивание же количества связей в полушариях при облучении свидетельствует об изменении степени согласованности различных показателей метаболизма в ЦНС, следствием которого может явиться нарушение пластических, энергетических и информационных составляющих системы ФМА.
Итак, исследование динамики уровня постоянного потенциала и микроэлементов в полушариях головного мозга мышей при нормальном и ускоренном старении показало, что облучение приводит к появлению в популяции большого количества животных с левой поворотной тенденцией; к снижению УПП и инверсии межполушарной разности потенциалов; к снижению концентрации микроэлементов в полушариях облученных животных; к перераспределению корреляционных связей между микроэлементами и выравниванию количества связей в полушариях головного мозга.
Отвечая на вопрос об устойчивости системы ФМА к неблагоприятным экстремальным воздействиям, необходимо отметить, что физиологически неравнозначные полушария обнаруживают различную чувствительность к радиационному повреждению: более чувствительным во всех экспериментах оказывалось доминантное полушарие, что приводило к нейрофизиологической, биохимической, а затем и к поведенческой инверсии ФМА.
Проблема устойчивости ФМА к неблагоприятному экстремальному воздействию, примером которого явилось радиационное повреждение ЦНС, также получила определенное разрешение: благодаря различной чувствительности к радиации физиологически неравнозначных полушарий, ФМА при радиационном воздействии претерпевает инверсию. Однако экстремальные неблагоприятные воздействия на организм имеют не только внешнее, техногенное, происхождение, но и внутренний, стрессогенный, генез. К таким воздействиям относятся патологические процессы, в частности, аутоиммунного генеза благодаря их системному характеру, вовлекающему в патологический процесс все жизненно важные системы, включая нервную и иммунную.
Изменению структурно-функциональной организации ФМА при развитии аутоиммунных системных ревматических заболеваний и ее роли в формировании иммунного ответа посвящена клиническая часть исследования.
Клинические исследования
Развитие нейроиммунологии в настоящее время привело к пониманию глубокой связи между нервной и иммунной системами. Изучению механизмов нейро-иммунной интеграции посвящены многочисленные исследования последних лет. Комплекс полученных результатов позволяет теперь рассматривать нервную и иммунную систему как единый структурно-функциональный блок, обладающий всеми признаками понятия "система" (включающего в себя три основных признака: структура, информация, управление). По определению фон Берталанфи, живая система представляет собой иерархически организованную открытую систему, сохраняющую себя или развивающуюся в направлении достижения состояния подвижного равновесия. Таким образом, нейро-иммунные взаимодействия в рамках единой системы выполняют в организме основные функции по адаптации к изменяющимся внешним (и внутренним) воздействиям (Н.Н. Брагина, Т.А. Доброхотова, 1981; В.В. Абрамов с соавт., 1991; 1996; А.В. Черноситов, 1995).
Многочисленные данные о сходстве фенотипических и функциональных параметров клеточных элементов иммунной и нервной систем, как и самих систем в целом, ставят вопрос об анализе закономерностей и основных принципов их взаимодействий. Поскольку ФМА является основным – интегрирующим – принципом организации ЦНС, ответ на поставленный вопрос невозможен без исследования роли ФМА в нейро-иммунных взаимодействиях. Существуют экспериментальные данные об изменении функциональной активности иммунокомпетентных клеток мышей в зависимости от повреждения того или иного полушария головного мозга. Р.Barneoud, (1990; 1993) показал, что мозговая кора модулирует иммунную систему асимметричным способом: экстирпация левосторонней коры влияет на иммунные параметры, в то время, как симметричная правая экстирпация или не влияет на иммунную систему или увеличивает ее активность. По мнению многих исследователей, нет никаких иммунных систем или гомеостатических механизмов защиты, которые в естественных условиях не подчинены взаимодействию между поведенческими и физиологическими событиями. В связи с этим традиционное представление о функциях иммунной системы, ограниченной клеточными взаимодействиями в пределах лимфоидных тканей, недостаточно, чтобы объяснить изменения в иммунитете, наблюдаемыми у животных и человека при реальных взаимодействиях организма и среды. Иммунная система может регулироваться, не только с помощью механизма обратной связи с нервными и эндокринными процессами, но с помощью механизмов прямой связи также. Результаты изучения механизма прямой связи иммунной системы свидетельствуют о том, что подобно прямым невральным и эндокринным воздействиям, поведение, при соответствующих обстоятельствах, может выполнять иммунорегуляторную функцию в естественных условиях.
В работе V.H. Denenberg (1991-а) крысы были подвергнуты двусторонним или односторонним токсическим повреждениям базальных ядер, в результате этого эксперимента было показано, что клетки базальных ядер включаются в комплексные взаимосвязи, существующие между ЦНС и иммунной системой.
Как известно, интерлейкины – естественно встречающиеся белки, которые регулируют и таким образом связывают иммунную систему и ЦНС, не проникая через гематоэнцефалический барьер. После инъекции меченого интерлейкина в мозг, в нем регистрировалась радиоактивность (с появлением меченого цитокина). Этот факт свидетельствует о наличии системы, которая переносит рекомбинант интерлейкина-альфа в неповрежденной форме от крови в ЦНС, осуществляя прямое соединение иммунной системы с ЦНС.
В работах С. Betancur (1991; 1992) показано, что мозг модулирует иммунную систему асимметричным способом. У мышей имеется зависимость между хиральностью и иммунной реакцией; также показано, что одностороннее удаление правого или левого полушария оказывает противоположное действие на некоторые параметры иммунной системы. Зависимость между продукцией аутоантител и предпочтением лапы наблюдалась у мышей-самцов, что ассоциировалось со связью между иммунной реактивностью, функциональной мозговой асимметрией и половой принадлежностью. У левшей обнаруживалось угнетение деятельности NK-клеток по сравнению с правшами. Левосторонние удаление коры подавляет действие NK-клеток, в то время как повреждения коры в правом полушарии не влияет на это показатель. P.J. Neveu и соавторами (1988; 1991; 1992) считают, что существует обратная связь между центральной нервной системой и иммунной системой. Активация иммунной системы приводит к выработке медиаторов; эти медиаторы активируют гипоталамус, который в свою очередь стимулирует выработку иммуносупрессивных молекул. В аутоиммунных процессах или других заболеваниях с чрезмерной активацией иммунной системы (например, рассматриваемые в этом разделе системные ревматические заболевания) в процессе стабилизации участвует гипоталамо-гипофизарно-надпочечниковая ось. Стресс является одной из причин снижения устойчивости к возбудителям инфекции и реактивации латентных вирусов. С другой стороны, стресс стимулирует продукцию цитокинов, которая в свою очередь может способствовать патогенезу воспалительных процессов неизвестной этиологии. Основываясь на том, что реакции иммунной системы включают ЦНС, гипоталамо-гипофизарно-надпочечниковую ось и иммунную систему, авторы исследовали влияние поведенческой латерализации на реакцию нервной, нейроэндокринной и иммунной системы к введению полисахарида LPS. Оказалось, что введение LPS вызвало улучшение мозговой деятельности благодаря увеличению норадренэргического, серотонинэргического и дофаминэргического обмена. Однако увеличение обмена серотонина, наблюдаемое в гипоталамусе и гиппокампе, произошло только в левом полушарии; кроме того, увеличение обмена серотонина в среднем гипоталамусе, повышение в плазме адренокортикотропина и уменьшение роста Т-лимфоцитов наблюдалось у мышей-правшей, но не у левшей. Таким образом, реакции иммунной системы выражены асимметрично и зависят от поведенческой латерализации.
C. Betancur и соавторы (1991) показали, что иммунитет мышей связан с латерализацией поведения: мыши-левши имели более низкую концентрацию IgM и IgG, чем мыши-правши. Авторы утверждают, что иммунные реакции животных, не отличающихся по возрасту, полу и генетическому фону связаны с функциональной асимметрией головного мозга, а гипоталамо-гипофизарно-надпочечниковая (ГГН) ось и симпатическая нервная система включаются в устойчивое соединение после инфекционной агрессии, инициируя асимметричный иммунный ответ. В серии экспериментальных исследований, проведенных P. Neveu с соавторами, показано, что центральная нервная система может модулировать иммунную систему, при этом иммуномдуляция может быть латерализованной. После односторонней экстирпации коры левого полушария у грызунов снижалась функциональная активность Т и В лимфоцитов, хотя те же функции оставались неизменными или даже возросли после удаления коры правого полушария. Тот же автор отмечает, что у людей с левосторонней моторной асимметрией (левшей) замечены нарушения в деятельности иммунной системы. С другой стороны, иммунная система может посылать информацию в ЦНС, которая кодируется и декодируется асимметрично. Эксперименты, проведенные P. Neveu (1992) на грызунах, показывают, что асимметричная мозговая модуляция может происходить также в подкорке с включением в нейроиммуномодуляцию дофамина. Таким образом, исследования, проводимые в течение многих лет этим автором, показали, что полушария мозга могут по-разному включаться в модуляцию иммунных реакций. Повреждения правой или левой стороны неокортекса приводит к противоположным изменениям различных иммунологических характеристик, таких, как пролиферация лимфоцитов, продукция интерлейкина-2, активация макрофагов или действие NK-клеток. Эти модели полезны для понимания иммуномодулирующих свойств мозга людей. Влияние межполушарной асимметрии на модуляцию иммунной системы можно наблюдать и на интактных животных: латерализация моторной асимметрии, то есть предпочтение лапы при пищевом подкреплении, коррелирует с реактивностью лимфоцитов, действием NK-клеток и продукцией аутоантител. Поэтому каждый биологический организм может быть охарактеризован типом латерализации вместе со специфическим типом иммунных и нейроэндокринных реакций. Эти межиндивидуальные различия могут быть ответственны за вариабельность среди членов популяции в реакциях на различные повреждающие воздействия, включая психологические стрессогенные факторы и инфекционные болезни. Кроме того, мозговая асимметрия проявляется в обмене веществ нейромедиатора и нейроэндокринной модуляции. Поскольку гиппокамп модулирует действие ГГН оси, авторы исследовали асимметрию распределения гиппокампально-кортикоидных рецепторов, предположив, что и эта асимметрия может различаться так же, как различается поведенческая латерализация животных. Результаты показывают, что независимо от поведенческой латерализации, имеется тенденция для правостороннего преобладания кортикоидных рецепторов в гиппокампе. Эти результаты свидетельствуют о том, что гиппокампальные рецепторы для кортикоидов могут играть важную роль в асимметричной мозговой модуляции иммунной системой.
Таковы теоретические предпосылки для исследования модулирующей роли функциональной межполушарной асимметрии при формировании иммунного ответа у людей, страдающих системными ревматическими заболеваниями.
По данным N. Geschwind (1984), заболевания иммунологической природы у леворуких людей встречаются в 2.5 раза чаще, чем у праворуких. Т.Я. Абрамова (1996) отмечает различия в иммунологических характеристиках у больных ревматоидным артритом с преимущественным повреждением суставов правой или левой стороны. Таким образом, данные разных авторов указывают на роль функциональной асимметрии полушарий головного мозга в патогенезе заболеваний, связанных с изменением функциональной активности иммунной системы. Однако, роль правого или левого полушария, как и всей системы межполушарной асимметрии в регуляции активности иммунных процессов нуждается в дальнейшем исследовании.
В клинических исследованиях на примере аутоиммунных заболеваний была проанализирована взаимосвязь между типом ФМА и иммунно-биохимическими показателями крови, характеризующими формирование иммунного ответа при ревматических заболеваниях. Сам факт наличия такой связи, как и достоверных различий в характеристиках иммунного ответа у больных с разным типом ФМА, является основанием для создания представления о модулирующей роли ФМА в формировании иммунного ответа при развитии системных ревматических заболеваний.
Поскольку инструментом для определения типа ФМА был УПП, т.е. его распределение в полушариях головного мозга, первый этап данного исследования был посвящен доказательству выдвинутого предположения о том, что постоянный потенциал является маркером аутоиммунного процесса. Исследования последних лет в области нейроиммунологии позволяют рассматривать аутоиммунные процессы как проявление дезинтеграции нервной и иммунной систем.
Одним из способов изучения взаимодействия центральной нервной и иммунной систем является анализ связей между иммунобиохимическими и нейрофизиологическими характеристиками. Аутоиммунные процессы сопровождаются явлениями энергодефицита и атрофии в ЦНС. В качестве показателя трофических и энергообменных процессов в ЦНС большой интерес вызывает изучение динамики УПП. Аутоиммунная агрессия сопровождается образованием иммунных комплексов, повреждающих эндотелий сосудов, и специфических антинейрональных антител. Таким образом, при развитии аутоиммунного процесса повреждаются все структуры, принимающие участие в формировании УПП. Системные ревматические заболевания в 80% случаев сопровождаются атрофическими процессами в ЦНС (В.А. Насонова, 1978; Р.Х. Кормейн с соавт., 1983; M. Bradbury, 1983; Bluestein H.G., 1988; Иванова М.М. с соавт., 1989; Я.А. Сигидин с соавт; 1994).
Клинические исследования были проведены на нескольких нозологических группах системных ревматических заболеваний, было обследовано 247 пациентов.
Больные находились на стационарном лечении в ревматологическом отделении КБ № 83, Москва, и получали одинаковую (гормональную) терапию. По нозологическим группам больные распределились следующим образом. Больных системной красной волчанкой (СКВ) было обследовано 109 человек, ревматоидным артритом (РА) – 102 человека, васкулитом (В) – 15, болезнью Бехтерева (ББ) – 11 и системной склеродермией (ССД) – 10 человек. Для статистической обработки результатов из всего контингента обследованных больных формировались группы по различным независимым критериям: нозологическим, нейрофизиологическим, по критерию остроты (активность), глубины и тяжести (стадия) патологического процесса и другим признакам. Возраст больных – от 15 до 64 лет. Группа сравнения была представлена 97 здоровыми донорами в возрасте от 18 до 60 лет. Для каждого больного был проведен общий клинический, биохимический и иммунологический анализ крови. Биохимические исследования касались трех видов обмена – белкового, липидного и углеводного, с широким спектром исследования ферментов. Электрофизиологические исследования (измерение и анализ УПП) были проведены у всех больных одновременно с клиническими, биохимическими и иммунологическими.
Кроме того, с целью исследования устойчивости структурно-функциональной организации межполушарной асимметрии при агрессивных экстремальных воздействиях на организм, было проведено электрофизиологическое обследование 13 человек – мужчин, участвовавших в 1986 г. в ликвидации аварии на ЧАЭС. Средний возраст обследованных составлял 50 ±1.2 лет. В качестве контроля были проведены измерения УПП у 13 доноров – мужчин такого же возраста, не имевших контакта с радиационным излучением.
Клинические методы определения диагноза, активности и стадии системных ревматических заболеваний проводилось на основе критериев Американской ревматологической ассоциации (1987), которые представляют собой список от 5 до 14 клинических признаков и данных инструментальных методов для каждого заболевания. Диагноз считается достоверным при наличии у больного определенного количества (от 3 до 7) этих критериев. Признак "активность" процесса был квалифицирован врачами-экспертами на основании как клинических, так и лабораторных проявлений заболевания с присвоением балла от 1 до 3 и основывался на совокупности биохимических и иммунологических показателей, свидетельствующих о тяжести заболевания.
Биохимические исследования были проведены по трем видам обмена – белковому, липидному и углеводному с широким спектром исследования ферментов. В основе всех исследований лежали унифицированные методики (Ф.И. Комаров с соавт., 1981). Всего было определено 32 клинических, биохимических и иммунологических показателя крови, представленных в таблице 3.
Таблица 3.
|
Клинические |
Hb, СОЭ, лейкоциты, лимфоциты, эритроциты, моноциты, сегментоядерные клетки, нейтрофилы |
|
Биохимические |
Серомукоид, b-ЛП, ОБ, a1, a2, b, g-глобулины, альбумин, АлАТ, АсАТ, КФК, ЩФ, ЛДГ, К+, Na+, глюкоза, мочевина, креатинин, мочевая кислота. |
|
Иммунологические |
СРБ, ЦИК, IgA, IgM, IgG |
Биохимические показатели сыворотки крови (общий белок, альбумин, креатинин, мочевую кислоту, α-1, α-2, ß- и γ-глобулины, ß-липопротеины, серомукоид, глюкозу, лактатдегидрогеназу (ЛДГ), креатинфосфокиназу (КФК), аспартатаминотрансферазу (АСТ), аланинаминотрансферазу (АЛТ), щелочную фосфатазу (ЩФ)) определяли на автоанализаторе Super-Z-818 фирмы Мицубиси (Япония). Концентрацию ионов натрия и калия в сыворотке измеряли на анализаторе Ciba Corning (Англия). Концентрации иммуноглобулинов IgA, IgM, IgG в сыворотке, уровень циркулирующих иммунных комплексов (ЦИК) и С-реактивного белка, компоненты ревматоидного фактора – лактекс-тест и реакция Ваалер-Розе были определены стандартными лабораторными методами, основанными на реакциях преципитации.
Активность компонентов сывороточного комплемента классического пути – С1, Сq, С2, С3, С4 и С5, а также общую гемолитическую активность классического пути СН50 определяли с использованием микропанелей для титрования и реагентов производства "Реаком" для определения индивидуальных компонентов комплемента по их гемолитической активности (Л.В. Козлов с соавторами, 1985; 1991).
УПП регистрировали с помощью аппаратно-программного диагностического комплекса “Нейроэнергон-01”– в котором используются методы анализа и топографического картирования УПП, что позволяет производить оценку суммарных энергозатрат головного мозга и его отдельных областей (В.Ф. Фокин с соавт., 1994).
Для регистрации УПП использовали неполяризуемые хлорсеребряные электроды, заполненные 30% раствором NaCl. Сопротивление электродов 10-20 кОм. Регистрация УПП производилась неинвазивно, непосредственно от кожи головы в пяти точках:- лобной (F), центральной (С), затылочной (О), правой (Тd) и левой (Тs) височных. Референтный электрод располагают на запястье с внутренней стороны.
После монополярной регистрации УПП головного мозга происходит компьютерный анализ распределения потенциала по поверхности головы с последующим картированием в соответствии со шкалой условных цветов. Заключительным этапом электрофизиологического обследования является сравнение УПП в различных областях мозга с последующим заключением о распределении энергозатрат в коре головного мозга пациента.
При вычислении множественных линейных коэффициентов корреляции применяли линейную пошаговую регрессию. В большинстве случаев значение F-включения признака в модель устанавливали равным 4, F-исключения – равным 3,9.
Начальный этап клинического исследования заключался в исследовании маркерной роли постоянных потенциалов в мониторинге развития патологического аутоиммунного процесса.
В данном исследовании исходили из представления о том, что информативными являются не только значения УПП в областях отведения (F, С, O, Тd и Тs), но и параметры, характеризующие взаимоотношения между УПП разной локализации. Для целей настоящей работы наибольший интерес представляет взаимосвязь межполушарной разности УПП между правым и левым височным отведением (Td-Ts) c совокупностью иммунологических и биохимических показателей.
Множественные линейные регрессии параметров УПП по иммунологическим и биохимическими пoкaзaтелям. С помощью процедуры пошаговой регрессии изучали связь между межполушарной разностью УПП и биохимическими и иммунологическими показателями. Таблица 4.
Величина R2 соответствует доле дисперсии зависимой переменной, объясненной линейной моделью. Включение всех признаков в модель высоко достоверно (p<0,005). Величина стандартизованного коэффициента регрессии (указана в скобках после каждого параметра в таблице) пропорциональна «вкладу» каждого предиктора в регрессию.
Таблица 4
|
Результаты множественной регрессии межполушарной разности УПП с иммунологическнм и биохимическим показателям
|
||||
|
№ |
Параметр УПП |
Иммунологические и биохимические показатели (в порядке вхождения в модель) |
R |
R2 |
|
9 |
(Td-Ts) |
ЛДГ (0,40), Hb (-1,5), мочевая кислота (0,56), ЩФ (-0,75), у-глобулин (-0,46), IgA (0,19), СРБ (-0,23), лимфоциты (-0,36), креатинин (-0,35) |
0,.97 |
0,95 |
Показатель межполушарной асимметрии (Td-Ts) имел высокую корреляцию с иммунобиохимическими показателями. Коэффициент множественной корреляции равен 0,97. Этот результат соответствует представлениям о модулирующей роли межполушарной асимметрии в организации иммунного ответа. Положительная связь (Td-Ts) с уровнем ЛДГ полностью соответствует показанной выше отрицательной связи между Ts и ЛДГ и нарастанию амбидекстрии при развитии повреждения клеток и тканей. Кроме того с этим показателем связана и концентрация IgA – специфическая характеристика для аутоиммунного процесса.
Аутоиммунная агрессия распространяется в направлении от крови к ткани мозга, затрагивая при этом все структуры, принимающие участие в генезе УПП. Иммунные комплексы повреждают стенки сосудов головного мозга. Проницаемость мембран эндотелиальных клеток увеличивается, их потенциал, по-видимому, уменьшается. Процессы повреждения распространяются на глиальные элементы, а затем и на нейроны. Все эти явления происходят несинхронно в разных полушариях головного мозга, что может приводить к увеличению разброса межполушарной разности УПП.
Полученные результаты дают основание для использования метода регистрации и анализа межполушарной разности УПП при изучении процессов интеграции нервной и иммунной систем, а также при мониторинге состояний организма, сопряженных с аутоиммунными процессами.
Глубина и характер повреждения ЦНС, а также скорость развития полиорганных нарушений при ревматических заболеваниях определяются, в первую очередь, активностью развивающегося аутоиммунного процесса. Задача мониторинга активности является актуальной как в фундаментальном, нейробиологическом, так и в практическом, медицинском, аспектах.
Традиционные методы нейрофизиологического исследования – ЭЭГ, компьютерная томография, вызванные потенциалы и др. – ориентированы либо на оценку процессов обработки информации в ЦНС, либо на регистрацию выраженных структурных изменений. Вероятно, поэтому они не всегда позволяют судить о характере и степени повреждения ЦНС и об активности аутоиммунного процесса.
Для этой цели оказался информативным интегральный нейрофизиологический показатель стационарных процессов в ЦНС – УПП. Поскольку он определяется потенциалами гематоэнцефалического барьера, глии и нейронов, т.е. именно тех структур, которые повреждаются при аутоиммунной агрессии, изменения УПП оказываются связанными с глубиной поражения ЦНС и активностью развивающегося аутоиммунного процесса.
В настоящем разделе исследования мы проанализировали связи между параметрами УПП и биохимическими и иммунологическими показателями крови, характеризующими активность аутоиммунного процесса. Методами статистического анализа показано, что независимо от нозологической формы активность процесса при СРЗ находит отражение в распределении УПП у конкретного больного.
На данном этапе были проанализированы данные 64-х больных с различной нозологией СРЗ (ББ-9 человек, РА-29, СКВ-18, ССД-8) в возрасте от 15 до 64 лет. Признак "активность" был квалифицирован врачами-экспертами с присвоением каждому клиническому наблюдению бала от 1 до 3-х.
Связь межполушарной разности УПП с активностью аутоиммунного процесса
Иммунологическое и биохимическое описание признака «активность».
Признак «активность» аутоиммунного процесса, определенный экспертами, связан с комплексом биохимических и иммунологических показателей, и энергетических показателей измеренных при данном обследовании. Эта связь была подтверждена в нашем исследовании с помощью метода линейного дискриминантного анализа.
Признак «активность» вошел в модели для межполушарной разности УПП (Td-Ts).
При построении регрессионной модели для межполушарной разности потенциалов (Td-Ts) наиболее значимым предиктором оказался показатель ЛДГ. Второе место занял признак «активность». Оба предиктора уменьшили исходную дисперсию межполушарной разности потенциалов примерно на 36%, что соответствует коэффициенту множественной корреляции R=0,60. При дальнейшем включении в модель показателей "лейкоциты", "серомукоид" и "латекс-тест" удалось объяснить 75% исходной дисперсии (Тd-Тs) (коэффициент множественной корреляции R=0,87.
Анализ регрессионного уравнения показал, что возрастанию активности аутоиммунного процесса соответствует выравнивание значений потенциалов во всех точках отведения: межполушарные различия сглаживаются. Подтверждением этого факта является прогрессивное изменение коэффициента латерализации при увеличении активности процесса. Определённый по формуле: К=2(Тd-Ts)/(Td+Ts) коэффициент межполушарной асимметрии распределения постоянного потенциала для первой активности он равен (–0,23), для второй(–0,06), для третьей он приобретает положительное значение и становится равным (0,01), таким образом, в терминальной стадии заболевания коэффициент межполушарной асимметрии по модулю в 20 раз меньше, чем в начальной (рис.12).

Рис. 12. Изменение коэффициента межполушарной асимметрии распределения постоянного потенциала в зависимости от активности СРЗ.
Таким образом, активность заболевания характеризуется закономерным изменением ФМА, что свидетельствует о непременном участии последней в формировании иммунного ответа при развитии патологического процесса. Подобные явления наблюдали ранее при тяжелых старческих деменциях, сопровождающихся резким снижением энергозатрат головного мозга (В.Ф. Фокин с соавт., 1994).
Изменение ФМА на разных стадиях ревматических заболеваний
Известно, что разной степени тяжести заболеваний соответствует определенная стадия синдрома неспецифической адаптации (Г. Селье, 1960; В.П. Леутин, Е.И. Николаева, 1988; Гаркави Л.Х. с соавт., 1990). Это относится и к аутоиммунным ревматическим заболеваниям: активный патологический процесс включает в себя и процесс неспецифической адаптации, влияющей на характер нейроимунных и межполушарных отношений. Эти изменения взаимно обусловлены и характер их взаимного влияния меняется в зависимости от стадии патологического процесса.
При формировании адаптивного ответа на экстремальные воздействия различной этиологии существенную роль играет функциональная межполушарная асимметрия (ФМА). Известно, что, кроме генетических предпосылок, ревматические заболевания могут возникать также из-за сбоев в работе иммунной системы, вызванных нарастающим стрессогенным влиянием неблагоприятных условий внешней среды. Характер межполушарных отношений, изменяющийся при этом, может служить критерием уровня адаптивных возможностей человека. Как было показано ранее, среди множества показателей, используемых в настоящее время для характеристики ФМА, выделяется своей репрезентативностью показатель интенсивности энергетических процессов в полушариях головного мозга – уровень постоянного потенциала (УПП). У здоровых людей в состоянии спокойного бодрствования значение УПП над височными областями доминантного полушария выше на несколько милливольт, чем над субдоминантным (В.Ф. Фокин, 1996 с соавт.).
Из предыдущего раздела ясно, что распределение УПП в коре больших полушарий головного мозга связано с активностью аутоиммунного процесса, влияющего на трофику ткани и состояние энергетического обмена в ЦНС. Можно предположить, что на разных стадиях ревматических заболеваний меняется вовлеченность правого или левого полушария в управление иммунными реакциями, изменяя при этом интенсивность энергозатрат в обоих полушариях и межполушарное распределение УПП. По мере углубления патологического процесса нарастает энергетическое истощение ЦНС, что может привести к выравниванию значений УПП в правом и левом полушариях.
На данном этапе работы были проанализировали результаты всего обследованного контингента больных СРЗ (247 человек) и всех нозологий: СКВ, РА, васкулит, ББ и ССД). Анализировали межполушарное распределение УПП. К типу ФМА с левополушарным доминированием по УПП относили пациентов, у которых (Td-Ts) < 2 мВ; к правополушарному типу с (Td-Ts) > 2 мВ; в остальных случаях считали, что доминирующее полушарие отсутствует.
У больных, находящихся в момент обследования на разных стадиях ревматического заболевания, исследовали динамику основного параметра, определяющего характер ФМА – межполушарную разность УПП в височных областях правого и левого полушарий (Td-Ts). Результаты исследования приведены на рисунке 13, из которого видно, что межполушарная разность потенциалов претерпевает достоверное изменение по мере развития заболевания. Левополушарное доминирование, наблюдаемое в группе сравнения и на начальной стадии СРЗ, постепенно сменяется правополушарным на второй и третьей стадиях. На четвертой стадии асимметричное распределение УПП в полушариях значительно снижается, сменяясь эквипотенциальными значениями УПП в обоих полушариях.
Детальное распределение полушарного доминирования в норме и на разных стадиях заболеваний приведено в таблице 5, эти данные соответствуют результатам, приведенным в предыдущем разделе, где показано нарастание эквипотенциальности коры полушарий при повышении активности ревматического процесса.
Таблица 5
Полушарное доминирование в норме и на разных стадиях системных ревматических заболеваний
|
|
Тип межполушарной асимметрии полушарий |
||
|
Стадии заболеваний |
Левополушарное доминирование |
Правополушарное доминированиев |
Эквипотенциальностьполушарий |
|
Нормаааа |
85 |
5 |
6 |
|
1 |
25 |
2 |
0 |
|
2 |
17 |
14 |
3 |
|
3 |
5 |
31 |
26 |
|
4 |
0 |
0 |
21 |
Примечание: цифры в таблице обозначают количество пациентов с тем или иным типом ФМА (из числа всех обследованных).
Исчезновение нейрофизиологического межполушарного градиента (по критерию УПП) рассматривается как неблагоприятный прогностический признак, свидетельствующий об истощении адаптивных возможностей ЦНС. Развитие неспецифической реакции адаптации при заболевании приводит к большему вовлечению правого полушария в управление иммунными процессами сначала за счет его активации; затем, по мере развития атрофических процессов в головном мозге, происходит снижение активности обоих полушарий до уровня их эквипотенциальности. По-видимому, левополушарное доминирование в большей степени, чем правополушарное, способно обеспечить надежную и экономичную (в смысле адаптационных резервов) резистентность организма к экстремальному воздействию. Этот вывод хорошо согласуется с работой N. Geschwind c соавторами (1984), в которой показано, что правши обладают более высоким иммунным статусом, чем левши.
Таким образом, можно заключить, что разным стадиям системных ревматических заболеваний соответствуют различные типы ФМА. На ранних стадиях преобладает левополушарное доминирование, на развернутых – правополушарное доминирование, терминальная стадия характеризуется потерей ФМА.
Модулирующая роль функциональной межполушарной асимметрии в формировании иммунного ответа при ревматических заболеваниях
Данные разных авторов указывают на роль асимметрии полушарий головного мозга в патогенезе заболеваний, связанных с изменением функциональной активности иммунной системы. Однако, роль правого или левого полушария, как и всей системы межполушарной асимметрии в регуляции активности иммунных процессов нуждается в дальнейшем исследовании. Была исследована взаимосвязь между типами межполушарных отношений и иммунно-биохимическими показателями крови, характеризующими формирование иммунного ответа при ревматических заболеваниях. Наличие такой связи, как и достоверных различий в характеристиках иммунного ответа у больных с разным типом ФМА, является основанием для создания представления о модулирующей роли ФМА в формировании иммунного ответа при развитии системных ревматических заболеваний.
Сравнение средних значений иммунно-биохимических показателей крови у правшей, левшей и амбидекстров было проведено с помощью t-критерия.
Анализировали данные обследования выборки из 47 больных - только ревматоидным артритом (РА) и системной красной волчанкой (СКВ). По результатам измерения УПП 20 человек были отнесены к левополушарным, 13 - к правополушарным, 14 - к эквипотенциальным.
Среднее арифметическое значение концентрации a2-глобулинов у правополушарных было достоверно ниже (р< 0.05), чем у эквипотенциальных. В то же время активность ЛДГ в крови правополушарных была значительно (p<0.01) выше, чем у левополушарных (таблица 6).
Таблица 6
Среднее значение биохимических показателей у больных СРЗ с разными типами функциональной асимметрии
|
Биохимические характеристики |
Левополушарные |
Правополушарные |
Эквипотенциальные |
|
Альфа(2)-глобулины |
8.2±0.38 |
7.4±0.33 |
9.06±0.46 |
|
Лактат дегидрогеназа |
290.7±11.9 |
438.08±60.7 |
324.07±15.3 |
В таблице указаны средние арифметические значения параметров и ошибки среднего. Выделены значения, имеющие достоверные различия (p<0,05) в пределах строки.
Известно, что a2-глобулины являются наиболее ярким показателем активности воспалительного процесса. Повышение их уровня у эквипотенциальных по сравнению с правополушарными соответствует результатам наших предшествующих исследований, согласно которым правополушарность и эквипотенциальность характеризуют развернутую и терминальную стадии заболевания (Л.Л. Клименко с соавторами, 1999), в то время как левополушарность соответствует начальной фазе заболевания (Рис.13.).

Рис. 13. Изменение межполушарной разности потенциалов (Td-Ts, мВ) на разных стадиях ревматических заболеваний. Н - контрольная группа.
Различия в активности ЛДГ у лево- и правополушарных - это, фактически, различия между начальной и развернутой стадиями заболевания. Очевидно, что повреждение тканей у правополушарных больных более выражено, чем у левополушарных, что отражается в различной активности ЛДГ.
При сравнении дисперсий иммунологических и биохимических показателей в обследованных группах различия наблюдались гораздо чаще, чем при сравнении средних арифметических значений. В таблице 7 показана встречаемость дисперсий параметров при сравнении больных, разделенных по нейрофизиологическому критерию (УПП) на группы с различным полушарным преобладанием УПП.
При сравнении левополушарных и правополушарных больных, дисперсии иммунно- биохимических параметров были достоверно больше у правополушарных (9 случаев) и лишь дисперсия уровня альбумина была больше у левополушарных. При сравнении левополушарных и эквипотенциальных, большие дисперсии чаще наблюдались у эквипотенциальных – 9 случаев, у левополушарных только для активности ЩФ и уровня креатинина дисперсии были больше, чем у эквипотенциальных. При сравнении правополушарных с эквипотенциальнымиами количество больших дисперсий было примерно одинаково.
Иммунно-биохимические показатели можно условно разделить на две части: характеристики активности воспалительного процесса (уровни иммуноглобулинов, концентрация белка, количество иммунокомпетентных клеток) и показатели повреждения тканей (активность тканевых ферментов в крови и концентрация продуктов белкового метаболизма). Как видно из таблицы 7, параметры, характеризующие активность, - первые 11 параметров - наиболее сильно варьируют в группах правополушарных и эквипотенциальных. Наиболее вариабельными по показателям деструкции тканей оказываются правополушарные, наименее - левополушарные.
Дисперсию показателя можно рассматривать как меру устойчивости системы. В этом случае повышение дисперсии может свидетельствовать о переходном, т.е. неустойчивом состоянии системы. В группе левополушарных, в которой представлены преимущественно больные на начальной стадии заболевания, изменения метаболизма еще невелики и касаются только альбумина - главного неспецифического транспортного белка организма.
В развернутой стадии, характеризующейся инверсией ФМА, неустойчивость захватывает большее число параметров организма и становится в этом смысле системной. Этому состоянию соответствует распространение повреждения тканей и эндотелия сосудов головного мозга в результате аутоиммунной агрессии. Различия в скорости развития процесса повреждения, переход основных иммунных и биохимических параметров в область “патологических” значений отражаются в нарастании дисперсии по этим показателям внутри группы правополушарных.
Терминальная стадия заболевания, ассоциированная с эквипотенциализацией коры полушарий (амбидекстрией) соответствует более устойчивому (чем в группе левшей) состоянию системы, но уже качественно иному, чем вначале, необратимо от него отличающемуся. Этот период СРЗ характеризуется генерализованной атрофией тканей, в том числе ЦНС, и иммунно - биохимические различия между больными уменьшаются по сравнению с левшами, хотя остаются все же больше, чем в группе правшей.
Таблица 7
Встречаемость различий дисперсий иммунно - биохимических показателей в группах левополушарных, правополушарных и эквипотенциальных
Буквами обозначены группы с достоверно (p<0,01, критерий F) большей дисперсией для каждой сравниваемой пары.
|
|
Сравниваемые группы |
||
|
Показатель |
Лево-правополушарные |
правополушарные – эквипотенциальные |
Левополушарные - эквипотенциальные |
|
Лимфоциты |
|
Э |
Э |
|
Моноциты |
П |
П |
|
|
СЯ-клетки |
|
|
Э |
|
Латекс-тест |
П |
П |
|
|
Тест реакции Ваалер-Розе |
|
Э |
Э |
|
IG A |
|
A |
|
|
IG M |
|
Э |
Э |
|
IG G |
П |
П |
|
|
Общий белок |
П |
|
Э |
|
Альбумин |
Л |
Э |
Э |
|
Альбумин/Глобулин |
|
Э |
|
|
AЛАТ |
П |
П |
Э |
|
АСАТ |
П |
|
Э |
|
КФК |
П |
|
Э |
|
ФЩ |
|
|
Л |
|
ЛДГ |
П |
П |
|
|
Мочевина |
П |
П |
|
|
Мочевая кислота |
|
П |
|
|
Калий |
П |
|
|
|
Креатинин |
П |
П |
Л |
П, Л, Э - правополушарные, левополушарные, эквипотенциальные, соответственно.
СЯ-клетки – сегментоядерные клетки, IG A – иммунноглобулин А, IG G – иммунноглобулин G, IG M – иммунноглобулин М, АЛАТ – аланинаминотрансфераза, АСАТ – аспартатаминотрансфераза, КФК – креатинфосфокиназа, ФЩ – щелочная фосфатаза, ЛДГ – лактатдегидрогеназа.
Для выявления связи ФМА и иммунно - биохимических показателей провели множественный регрессионный анализ. Зависимой переменной взяли межполушарную разность потенциалов (Td-Ts), независимыми - все измеренные в работе иммунологические и биохимические параметры. В результате была получена функция регрессии, включающая показатели (в порядке вхождения в модель): активность ЛДГ, общий белок, число лейкоцитов, концентрация гемоглобина, активность КФК, уровень мочевой кислоты, активность щелочной фосфатазы, концентрация эритроцитов и С-реактивного белка:
(Td-Ts)рассч = 0,40 ЛДГ - 1,5 Hb + 0,56 Моч. к-та - 0,75 ЩФ - 0,46 g-глобулин + 0,19 IgA - 0,23 СРБ - 0,36 Лимфоциты - 0,35 Креатинин
R = 0,97; R2=0,95
Таким образом, тип ФМА, может быть определен и через комплекс иммунно-биохимических характеристик. Этот факт служит подтверждением модулирующей роли ФМА в формировании иммунного ответа.
Структурно-функциональная организация межполушарной асимметрии при радиационном облучении. Особенности медленной электрической активности головного мозга у ликвидаторов аварии на Чернобыльской АЭС
Нейрофизиологические паттерны при повреждающем действии ионизирующего облучения на организм носят многоуровневый характер. Радиационное поражение головного мозга вызывает структурные, нейрохимические, сосудистые, биоэлектрические, энергетические и многие другие изменения, приводящие в конечном итоге к атрофическим процессам и полному истощению ЦНС.
В ряде клинических и экспериментальных исследований показано, что биоэлектрическая активность головного мозга чрезвычайно чувствительна к радиационному воздействию: зарегистрировано снижение амплитуды вызванных потенциалов, замедление ритмов ЭЭГ и появление судорожных пиковых разрядов (W.L. McFarland et al., 1974; Б.И. Давыдов и др., 1991; E.M. Caparelli-Draquer et al., 1991; Л.К. Обухова и др., 1997).
Облучение вызывает падение артериального давления и снижение церебрального кровотока, что в свою очередь сопровождается снижением температуры мозга. Радиационное воздействие вызывает деструктивные процессы в барьерных механизмах мозга (А.Д. Рева, 1974; Доклад ООН, 1982; Ушаков И.Б. и др.,1987). Возрастание свободнорадикального окисления одновременно со снижением активности ферментов защиты – супероксиддисмутазы и системы глутатионовых ферментов – приводит к увеличению проницаемости мембран нейронов и глии и изменению их электрогенных свойств (В.Ф. Фокин и др., 1989; В.А. Барабой, 1991; De la Torre, 1998). Увеличение проницаемости гематоэнцефалического барьера тесно связано с радиационным повреждением эндотелия капилляров ( Cockerman et al., 1986; Ушаков и др., 1987).
Облучение вызывает глубокие нарушения энергетического метаболизма в ЦНС, сопровождаемые структурными изменениями в митохондриях нервных клеток, разобщением окислительного фосфорилирования и снижением концентрации АТФ в мозге (Е.Pausescu et al., 1974; О.В. Протасова и др., 1997; F. Van Bell, 1998).
Исследования последних лет позволяют рассматривать постоянный потенциал как удобный инструмент для исследования механизма пострадиационного изменения энергетического метаболизма ЦНС (Л.Л.Клименко с соавт., 1999 ).
На данном этапе работы мы исследовали пострадиационные изменения ФМА и энергетического метаболизма ЦНС у ликвидаторов аварии на ЧАЭС с помощью регистрации и анализа медленной электрической активности головного мозга.
Было обследовано 13 человек – мужчин, средний возраст которых составлял 50 ±1.2 лет, участвовавших в 1986 г. в ликвидации аварии на ЧАЭС. В качестве контроля были проведены измерения УПП у 13 доноров – мужчин такого же возраста, не имевших контакта с радиационным излучением. УПП регистрировали согласно описанной ранее методике. неинвазивно от кожи головы с помощью аппаратно-программного диагностического комплекса "Нейроэнергон-01".
Особенно важным в этом исследовании является факт достоверного различия показателя межполушарной асимметрии – разности потенциалов в правой и левой височных областях (Td – Ts): у ликвидаторов этот показатель в 4 раза выше, чем у доноров, что связано с достоверно более низким, чем у доноров, постоянным потенциалом в правой височной области (таблица 8)
Таблица 8
Средние значения показателей постоянного потенциала коры головного мозга у доноров и ликвидаторов последствий аварии на ЧАЭС
|
Параметры УПП |
Доноры |
Ликвидаторы |
|
F |
4.03 ±1.40 |
3.08±1.37 |
|
C |
7.85±1.06 |
9.44±1.56 |
|
O |
4.67±2.15 |
3.12±1.54 |
|
Td |
5.45±1.09* |
2.00±1.11* |
|
Ts |
6.12±1.80 |
4.67±1.82 |
|
Td-Ts |
-0.67±0.55* |
-2.66±1.05* |
![]()
![]()
* отмечены достоверно различающиеся значения (р<0,05)
Известно, что правое полушарие мозга больше, чем левое, связано с вегетативными функциями организма и меняет свою биоэлектрическую активность при стрессе различного происхождения (О.С. Адрианов, 1976; Н.Н. Брагина и др., 1981; В.Л. Бианки, 1989). Экстремальное повреждающее воздействие радиации приводит к накоплению токсичных продуктов свободнорадикального окисления, играющих роль неспецифического первичного медиатора радиационного стресса (В.А. Барабой и др., 1992). Адаптация к неблагоприятным воздействиям внешней среды осуществляется под контролем центральной нервной системы; очевидно, что управление многими измененными вегетативными функциями в ходе развития стресс-реализующих реакций происходит на фоне снижения медленной электрической активности различных областей коры головного мозга, в том числе и правого полушария. При сохранении близкого к норме значения постоянного потенциала в левом полушарии высокий показатель межполушарной асимметрии (Td – Ts) у ликвидаторов может быть как показателем высокой пластичности ЦНС, так и отражением мобилизации адаптивных резервов организма в условиях стресса, вызванного облучением (известно, что межполушарная асимметрия является показателем устойчивости ЦНС) (Н.Н. Брагина с соавт., 1981; В.Л Бианки, 1985; 1989; А.В. Черноситов, 1995).
При исследовании нейрофизиологических паттернов как у людей, подвергшихся радиационному облучению в результате аварии на ЧАЭС, так и у здоровых доноров, было показано достоверное различие средних значений постоянного потенциала, зарегистрированного в разных областях мозга. Во всех отведениях, кроме центрального, показатели постоянного потенциала у ликвидаторов были ниже, чем у доноров, что свидетельствует о пострадиационном энергодефиците мозга.
Итак, в результате проведенного исследования удалось ответить на заявленные во вступлении вопросы. ФМА – это многоуровневая иерархическая система, внутри которой предыдущий уровень рассматривается как механизм для последующего, более высоко расположенного уровня. Нейрофизиологические, биохимические, энергетические и поведенческие корреляты ФМА находятся между собой в динамическом взаимодействии, их возрастная динамика синхронизирована и взаимообусловлена, а структура связей внутри системы меняется в пространстве и времени. Именно система связей обеспечивает структуру ФМА, поддержание которой требует энергозатрат, меняющихся в процессе старения: энергозатраты максимальны в репродуктивный период, когда процесс выбора ассоциируется с выполнением генетической программы продолжения вида. ФМА – устойчивая система, она сохраняется и при старении, и при агрессивном радиационном воздействии, однако полушария стареют с разной скоростью и обладают разной чувствительностью к радиационному облучению.
При вовлеченности организма в стресс-реализующий патологический процесс ФМА принимает участие в модуляции иммунного ответа, закономерно изменяясь с развитием аутоиммунного заболевания. Характеризующая ФМА межполушарная разность УПП является значимым предиктором иммунного ответа и, соответственно, активности и стадии аутоиммунного процесса. При генерализованной аутоиммунной агрессии, распространяющейся в направлении от крови к ткани мозга, затрагиваются все компоненты, принимающие участие в генезе УПП. Иммунные комплексы повреждают стенки сосудов головного мозга, проницаемость мембран эндотелиальных клеток увеличивается, их потенциал уменьшается, процессы повреждения распространяются на глиальные элементы, а затем и на нейроны. Все эти явления происходят несинхронно в разных отделах головного мозга, что вначале приводит к увеличению разброса УПП. Однако при распространении повреждений и расширении очагов атрофии на значительные области головного мозга разнообразие сокращается вследствие тотального повреждения мембран клеток нервной системы. Результатом развития этого драматического сценария становится эквипотенциализация коры мозга, дезинтеграция нервной и иммунной систем, утеря асимметричной организации.
При повреждающем радиационном воздействии ФМА сохраняется, но динамически изменяется вместе с изменением энергообмена. Эти изменения сохраняются и в отдаленные сроки после облучения: спустя 12 лет после аварии на ЧАЭС мозг ликвидаторов «помнит» о полученном в 1986 году радиационном поражении, демонстрируя пострадиационный энергодефицит, деструкцию межцентральных отношений и, соответственно, ухудшение процессов передачи информации в ЦНС.
Таким образом, все многообразие изменений ФМА и сопряженных с ними изменений иммуно-биохимического статуса организма в конечном итоге служит для оптимизации адаптивных процессов при стресс-реализующем патологическом процессе внешнего или внутреннего генеза. Отводя ЦНС традиционно ведущую роль в процессах управления, можно утверждать, что структурно-функциональная организация межполушарной асимметрии подчинена задаче мобилизации и оптимизации многоуровневого каскада адаптивных реакций, осуществляя управление формированием адаптивными реакциями в пространственно-временном континууме внешних и внутренних воздействий на организм как открытую систему.
Литература
1. Абрамов В.В. Интеграция иммунной и нервной систем. Новосибирск: Наука, 1991. С. 167.
2. Абрамов В.В., Абрамова Т.Я. Асимметрия нервной, эндокринной и иммунной систем. Новосибирск: Наука, 1996. С. 97.
3. Адрианов О.С. О принципах организации интегративной деятельности мозга.-М.:Медицина, 1976.-279 с.
4. Анисимов В.Н. Солнечные часы старения // Природа.- 1995.- № 10.- С. 3 - 12.
5. Афифи А., Эйзен С. Статистический анализ: подход с использованием ЭВМ. М.: Мир. 1982. 488 с.
6. Барабой В.А., Брехман И.И., Глотин В.Г., Кудряшов Ю.Б. Перекисное окисление и стресс. - Санкт- Петербург: Наука. - 1992. -148 с.
7. Барабой В.А.,Орел В.Э., Карнаух И.М. Перекисное окисление и радиация. - Киев: Наукова думка. - 1991. - 256 с.
8. Бианки В.Л. Механизмы парного мозга - Л.: Наука, 1989.- 264с.
9. Биохимия мозга. Под ред И.П. Ашмарина. –С-Пб. 1999. 325 с.
10. Брагина Н.Н., Доброхотова Т.А. Функциональная асимметрия человека. - М.: Медицина. -1981.-288 с.
11. Гаркави Л.Х., Квакша Е.Б., Уколова М.А. Адаптационные реакции и резистентность организма. Ростов-на-Дону: Изд-во Рост. ун-та, 1990. С. 222.
12. Давыдов Б.И., Ушаков И. Б., Федоров В.П. Радиационное поражение головного мозга. - М.: Энергоатомиздат. - 1991. - 240 с.
13. Делъва В.А. Содержание и топография некоторых микроэлементов в головном мозгу человека в норме и патологии. Автореф. дисс.... док. мед. наук. Киев. 1965.
14. Диксон М., Уэбб Э. Ферменты М: Мир. ТТ 1, 2, 3 1982.
15. Дудел Дж., Рюэтт И., Шмидт Р., Янг В. Физиология человека. М.: Мир, 1985. Т.1. С.228.
16. Иванов К.П. Биоэнергетика и температурный гомеостазис. - Л.: Наука. - 1972 - 171с.
17. Иванова М.М., Близнюк О.И., Тодуа Ф.И. Поражение центральной нервной системы при системной красной волчанке по данным компьютерной томографии // Клин. Мед. – 1989. – №12. – С. 93-98.
18. Илюхина В.А., Заболотских И.В. Энергодефицитные состояния здорового и больного человека. - С - Пб.: Наука, 1993. - 192 с.
19. Ионизирующее излучение: источники и биологические эффекты. Нью-Йорк: ООН. 1982. 779 с.
20. Карнаухов В.Н. Функции каротиноидов в клетках животных. М.: Наука, 1973. 104 с.
21. Клименко Л.Л. Динамика показателей энергетического метаболизма коры больших полушарий головного мозга в позднем онтогенезе крыс. // Известия АН. Сер. Биол. – 2001. – №2. – С. 213-219.
22. Клименко Л.Л. Многоуровневая организация функциональной межполушарной моторной асимметрии // Биофизика. 1987. Т. 32. В. 4. С. 696.
23. Клименко Л.Л. Температурная асимметрия больших полушарий головного мозга крыс. // Известия АН. Сер. Биол. – 2000. – №6. – С. 721-727.
24. Клименко Л.Л. Фокин В.Ф., Деев А.И., Конрадов А.А. Динамика корреляционных связей между уровнями системы межполушарной асимметрии при старении крыс // Цитология-1994.-Т. 36, № 7.-С. 738.
25. Клименко Л.Л., Грызунов Ю.А., Кудряшова Л.П., Турна А.А., Матвеев Г.Н., Протасова О.В., Максимова И.А., Комарова М.Н., Фокин В.Ф. Активность аутоиммунного процесса при системных ревматических заболеваниях и распределение постоянного потенциала головного мозга // Биофизика.– 1999 – Т. 44, вып. 2.– С. 368-371.
26. Клименко Л.Л., Грызунов Ю.А., Кудряшова Л.П., Турна А.А., Матвеев Г.Н., Протасова О.В., Максимова И.А.. Комарова М.Н., Фокин В.Ф. Постоянные потенциалы головного мозга как нейрофизиологические маркеры аутоиммунного процесса // Биофизика.– 1999.- Т. 44, вып. 2.– С. 372-377.
27. Клименко Л.Л., Деев А.И., Протасова О.В., Конрадов А.А., Фокин В.Ф., Пирузян Л.А. Синхронизация изменений уровня постоянного потенциала и концентрации продуктов перекисного окисления липидов головного мозга в онтогенезе у крыс.// Биофизика. – 1999. – Т.44, вып. 3. – С. 540-544.
28. Клименко Л.Л., Деев А.И., Протасова О.В., Конрадов А.А., Фокин В.Ф. Системная организация функциональной межполушарной асимметрии. Зеркало асимметрии. // Биофизика. – 1999. – Т.44, вып. 5. – С. 916-920.
29. Клименко Л.Л., Деев А.И., Протасова О.В., Фокин В.Ф. Системная организация функциональной межполушарной асимметрии и ее изменение в позднем онтогенезе крыс.// Успехи геронтологии. – 1998. – выпуск 2. – С. 82-89.
30. Клименко Л.Л., Деев А.И., Протасова О.В., Фокин В.Ф., Пирузян Л.А. Динамика функциональной межполушарной асимметрии и половой диморфизм в онтогенезе у крыс. // Биофизика. – 1999. – Т.44, вып. 5. – С. 921-922.
31. Клименко Л., Деев А., Протасова О., Фокин В., Пирузян Л. Асимметрия старения больших полушарий головного мозга крыс.//Биофизика.– 1998 – Т. 43. – Вып. 6.– С. 1063-1065.
32. Клименко Л.Л., Деев А.И., Фокин В.Ф. Корреляция содержания флуоресцирующих пигментов старения в гомогенатах коры мозга крыс с уровнем постоянного потенциала мозга в позднем онтогенезе // Биофизика.- 1987.-№ 4.-С.691-692.
33. Клименко Л.Л., Козлов Л.В., Кудряшова Л.П., Протасова О.В., Максимова И.А., Комарова М.Н., Конрадов А.А., Турна А.А., Союстова Е.Л., Матвеев Г.Н., Фокин В.Ф. Физические критерии патологических процессов. III. Структура корреляционной связи уровня постоянных потенциалов головного мозга с белками сывороточного комплемента при ревматических заболеваниях // Биофизика.– 1999.- Т. 44, вып.1.– С. 128-131.
34. Клименко Л.Л., Коломина Л.Н., Фокин В.Ф. Связь параметров перекисного окисления липидов с уровнем постоянного потенциала головного мозга крыс // Биофизика.- 1987.-№4.-С.689-б90.
35. Клименко Л.Л., Кудряшова Л.П., Протасова О.В., Максимова И.А., Комарова М.Н., Конрадов А.А., Матвеев Г.Н., Фокин В.Ф. Физические критерии патологических процессов. I. Изменение асимметрии функциональной межполушарной на разных стадиях ревматических заболеваний // Биофизика.– 1999.- Т. 44, вып.3.– С. 537-539.
36. Клименко Л.Л., Кудряшова Л.П., Протасова О.В., Максимова И.А., Комарова М.Н., Союстова Е.Л., Турна А.А., Матвеев Г.Н., Фокин В.Ф. Физические критерии патологических процессов. II. Модулирующая роль функциональной межполушарной асимметрии в формировании иммунного ответа при ревматических заболеваниях // Биофизика.– 1998.- Т. 43, вып.6.– С. 1066-1070.
37. Клименко Л.Л., Обухова Л.К., Деев А.И., Протасова О.В., Комарова М.Н., Фокин В.Ф., Пирузян Л.А. Нейрофизиологические и биохимические паттерны функциональной межполушарной асимметрии мышей при нормальном и ускоренном старении. // Клиническая геронтология. – Т.7.– №5-6. С. 30-34.
38. Клименко Л.Л., Протасова О. В., Конрадов А. А., Фокин В.Ф. и др. Микроэлементы в нейрофизиологических механизмах формирования межполушарной асиметрии // Известия РАН. Сер. биологическая. -1995.-№ 4.-С. 422-429.
39. Клименко Л.Л., Протасова О.В, Обухова Л.К., Комарова М.Н., Пирузян Л.А. Динамика уровня постоянного потенциала и микроэлементов в полушариях головного мозга мышей при нормальном и ускоренном старении. // Клиническая геронтология. – Т.7.– №7. С. 23-28.
40. Клименко Л.Л., Протасова О.В., Конрадов А.А, Фокин В.Ф. Пирузян Л.А. Микроэлементы в нейрофизиологических механизмах формирования межполушарной асиметрии // Известия РАН. Сер. биологическая. -1995.-№ 4.-С. 422-429.
41. Клименко Л.Л., Протасова О.В., Максимова И.А., Турна А.А., Матвеев Г.Н., Союстова Е.Л., Комарова М.Н., Фокин В.Ф. Особенности медленной электрической активности головного мозга у ликвидаторов аварии на Чернобыльской АЭС // Известия РАН, сер. Биол. – 1999. – №3. – С. 337-342.
42. Клименко Л.Л., Фокин В.Ф., Деев А.И., Конрадов А.А. Динамика корреляционных связей между уровнями системы межполушарной асимметрии при старении крыс // Цитология-1994.-Т. 36, № 7.-С. 738.
43. Козлов Л.В., Агеев В.П., Сизой М.Н., Мельник Е.И., Дедов И.И., Шамхалова М.Ш., Абугова И.А. Инсулинзависимая активность комплемента в сыворотке больных сахарным диабетом. // Клиническая медицина. – 1991. – №3. – С.60-64.
44. Козлов Л.В., Вавилова Л.М., Голосова Т.В. Микрометод определения факторов комплемента. // Иммунология. –1985. – №3. – С. 66-68.
45. Комаров Ф.И., Коровкин Б.Ф., Меньшиков В.В. Биохимические исследования в клинике. М.: Медицина. 1981. 406 с.
46. Кормейн Р.Х., Асгар С.С. Иммунология и болезни кожи. М.: Медицина, 1983. 254 с.
47. Леутин В.П., Николаева Е.И. Психофизиологические механизмы адаптации и функциональная асимметрия мозга. Новосибирск: наука. 1988. 192 с.
48. Насонова В.А, Соловьев С.К, Иванова М. Клиническая ревматология. М.М Медицина. 1989.
49. Ноздрюхина Л.Р. Биологическая роль микроэлементов в организме животных и человека. М.: Наука. 1977. 183 с.
50. Обухова Л.К, Эмануэль Н.М. Роль свободнорадикальных реакций окисления в молекулярных механизмах старения живых организмов. // Успехи химии. –1983.– Т. 52, № 3. –С. 353-372.
51. Обухова Л.К. Свободнорадикальные механизмы старения в биологической эволюции // Итоги науки и техники. Общие проблемы биологии- 1986.- Т. 5.- С. 36-68.
52. Обухова Л.К., Клименко Л.Л., Соловьева А. С. Изменение функциональной моторной асимметрии при старении и после облучения // Известия РАН. Сер. биологическая.-1997.-№ З.-С. 315-319.
53. Пасечник В.И., Кузнецова Г.Д. Оценка теплопродукции, вызываемой волной распространяющейся депрессии // Биофизика.-1987.- Т. 32, № l.-C. 145-149.
54. Д. Пельц, Ф. Эндрюс Ученые в организациях. Об оптимальных условиях для исследований и разработок. М.: Прогресс, 1973. 471 с.
55. Потапенко А.И., Обухова Л.К. Дефекты вторичной структуры ДНК, опознаваемые нуклеазой S1, и их возможная роль в старении соматических клеток млекопитающих. // Известия Ран. Сер. Биол.– 1992. – №6.- С. 940
56. Протасова О.В., Максимова И.А., Клименко Л.Л., Турна А.А., Конрадов А.А. Отдаленные последствия действия облучения при ликвидации последствий Чернобыльской катастрофы на концентрацию макро- и микроэлементов в сыворотке крови человека. // Известия АН. Сер биол. – 1997. – №5. – С. 592-595.
57. Рабек Я. Экспериментальные методы в фотохимии и фотофизике. М.: Мир, 1985 С.799-801.
58. Райцес B.C. Нейрофизиологические основы действия микроэлементов. Л. 1981. С. 149.
59. Рева А.Д. Ионизирующее излучение и нейрохимия. М.: Атомиздат, 1974. 274 с.
60. Рунион Р. Справочник по непараметрической статистике: англ. - М: Финансы и статистика, 1982 - 197 с.
61. Селье Г. Очерки об адаптации. М.: Медгиз. 1960. 251 с.
62. Сигидин Я.А., Гусева Н.Г., Иванова М.М. Диффузные болезни соединительной ткани. М. Медицина, 1994. 541 с.
63. Справочник по ревматологии. Под ред. В.А. Насоновой. М.: Медицина, 1978. 256 с.
64. Ушаков И.Б., Карпов В.Н. Изменение проницаемости гематоэнцефалического барьера при комбинированном воздействии Y -облучения и измененной газовой среды // Радиобиология. - 1987. - Т. 33, В. 2 - С. 839-841.
65. Фокин В.Ф. Центрально - периферическая организация функциональной моторной асимметрии: Дис. д-ра биол. наук: 03.00.13 М., 1982. 460 с.
66. Фокин В.Ф., Пономарева Н.В., Андросова Л.В., Секирина Т.П, Нейроимунные отношения при нормальном старении и деменциях Альцгеймеровского типа // Вестник РАМН.- 1996.- №3 - 4.- С. 32 - 39.
67. Фокин В.Ф., Пономарева Н.В., Букатина Е.Е. Уровень постоянного потенциала головного мозга человека в молодом, зрелом и старческом возрасте // Патол. физиол. и экспер. терапия,-1986.- №6.-С.-72-74.
68. Фокин В.Ф., Пономарева Н.В., Гаврилова С.И. Энергетичекий аспект деятельности головного мозга при нормальном старении и болезни Адьцгеймера // Вестник РАМН.- 1994.-№ 1.-С. 39-42.
69. Фокин В.Ф., Пономарева Н.В., Орлов О.Н. и др. Связь электрических реакций головного мозга с процессами перекисного окисления липидов при патологическом старении // Бюлл. экспер. биологии..-1989.- № 6.- С.682-684.
70. Черноситов А. В. Неспецифическая резистентность к экстремальным воздействиям в зависимости от характера функциональных межполушарных отношений: Дис .. д-ра биол. наук. Ростов-на-Дону, 1995. 255 с.
71. Шевелев И.А., Кузнецова Г.Д., Цыкалов Е.Н., Горбач А.М. и др. Термоэнцефалоскопия.- М.: Наука, 1989.- 223 с.
72. Age pigments / Ed. by R.S. Sohal. North Holland: Elseiver, Amsterdam: Biomedical Press, N.Y.: Oxford, 1981. 394 p.
73. Barneod P., Van der Loos H. Direction of handedness linked to hereditary asymmetry of a sensory system // Proc. Nat. Acad. Sci. USA. 1993. V. 90. № 8. P. 3246.
75. Betancur C., Sandi C., Vitiello S. et al. Activity of the hypothalamo-pituitary-adrenal axis in mice selected for left- or right-handedness // Brain Res. 1992. V. 589. P. 302.
76. Bjorkerud S. Isolated lipofuscin granules a survey of a new field. // Advances Gerontol. Res.– 1964.– V. 1.- P. 257.
77. Black M.S., Thomas M.B., Browner R.F. Determination of metal chelates byinnductivety coupled plasma atomic emission spectrometry and application to biological materials //Anal. Chem. 1981. V. 53. № 14. Р. 2224.
78. Bluestein H.G. Nervous system disease in systemic lupus erythematosus// Immunol. Allerg. Clin. N. Amer. – 1988. – V.8., № 2. – Р. 315-329.
79. Bradbury М. The concept of a Blood - Brain Barrier-N-Y-1983.- 479 P.
80. Bures J., Buresova 0., Krivanek J. The Mechanism and Application of Leao Spreading Depression of Elec-troencephalographic Activity. Prague: Acad. Press, 1974.
81. Busto R, Dietrich W.D., Glob. us M.Y. et al. Small differences in intraischemic brain temperature critically determine the extent of ischemic neuronal injury // J. Cereb. Blood Flow Metab-1987-V. 7-№ 6-P. 729-738. - 1990. - V. 15. - N8. - Р. 815 - 820.
82. Busto R.,Dietrich W.D., Globus M.Y. et al. Small differences in intraischemic brain temperature critically determine the extentt ofischemic neuronal injury // J. Cereb. Blood Flow Metab.-1987-V. 7.-№ б-Р. 729-738.
83. Cain D.P., Hargreaves E.L., Boon F. Brain temperature- and behavior-related changes in the dentate gyms field potential during sleep, cold water immersion, radiant heating, and urethane anesthesia / /Brain Res.- 1994-V. 658.-N 1-2-P. 135-144.
84. Caparelli -Draquer E.M., Schmidt S.L. Abscence of population cerebral asymmetries in mice with callosal defects induced by prenatal gamma irradiation // Braz. J. Med. Biol. Res. - 1991. -V. 24. -P. 1233 - 1238.
85. Cеnrасес J. Interaction of the cerebral hemispheres in relation to their functional asymmetry // Bratislav. Lek Listy. - 1991 - V. 92. - P. 515 - 520.
86. Cockerman L.G., Cerveny T.J., Hampton J.D. Postradiation regional cerebral blood flow in primates // Aviat. Space a nd Environ. Med. - 1986. - V. 57, N 6. -Р. -578 - 582.
87. Collins R.L. On the inheritance of direction and degree of asymmetry. In: Cerebral lateralization in nonhuman species. Ed. Glick S.D. N.Y.: Acad.Press; 1985. P.41.
88. Collins R.L. On the inheritance of handedness. I. Laterality in inbred mice // J. Hered. 1968. V. 59. № 1. P. 9.
89. Collins R.L. Reimpressed selective breeding for lateralization of handedness in mice // Brain Res. 1991. V. 564. № 2. P. 194.
90. Collins R When left-handed mice live in right-handed worlds//Science. 1975 V.187 №4172 P.181.
91. De la Torre J.C., Nelson N., Sutherland R.J., Pappas B.A. Reversal of ischemic-induced chronic memory a free radical scavenger-glycolytic intermediate combination // Brain Res .-1998—V. 777, N 1-P. 285-288.
92. Denenberg V. Evolution proposes and ontogeny disposes // Brain Lang. 2000. V. 73. № 2. P. 274.
93. Denenberg V.H. Hemispheric laterality in animals and the effects of early experience // Behav. Brain Sci. 1981. V. 4. № 1. P. 1.
94. Denenberg V., Mobraaten L., Sherman G. et al. Effects of autoimmune uterine/maternal environment on cortical ectopias, behavior and autoimmunity//Brain Res. 1991a.V.563. №1. P. 114.
95. Denenberg V.H., Sherman G.F., Morrison L. et al. Behavior, ectopias and immunity in BD/DB reciprocal crosses // Brain Res. 1992. V. 571. № 2. P. 323.
96. Denenberg V.H., Sherman G.F., Schrott L.M. et al. Spatial learning, discrimination learning, paw preference and neocortical ectopias in two autoimmune strains of mice // Brain Res.1991b. V. 562. № 1. P. 98.
97. Fokin V.F., Ponomareva N. V., Klimenko L.L. Dynamiks of catabolic processes in the human brain and development of psychopathologic reactions in aging and degenerative diseases // Behavioural Pharmacology.-1995.-Vol. 6, Suppl. l.-P. 54-55.
98. Geschwind N,. Behan P. Hormones,handedness and immunity // Immunol. Today.- 1984. – V. 5, №7.– Р.190-191.
99. Glick S.D., Jerussi T.P., Zimmerberg B. Behavioral and neuropharmacological correlates of nigrostriatal asymmetry in rats. In: Lateralization in the Nervous System. Eds. Harnad S., Doty R.W., Goldstein L. et al. 1977. N.Y.: Acad.Press. P.213.
100. Gumming G.M. The copper and iron cotent of brain and liver in the normal and hepato-lenticular degeneration // Brain. 1948. V. 71. №1-4. P. 109.
101. Gumnit R DS shifts accompanying seizure activity // Handbook of Amsterdam, Elsevier.- 1974a:-V.10, part. A.-P. 66-76..
102. Gumnit R. Field potentials in partial seizures.-A reppraisal // Origin of fields potentials: Intern Symposium, Muenster, Germany.-Stuttgart.-1979-P. 183-194.
103. Hansen A.J. Effect of anoxia on ion distribution in the brain // Phisiol. Rev. – 1985. – V. 65, N1. – P.101-148.
104. Hodgkin A.L. The conduction of the nervous impulse // Liverpool: Univ. Press.-1964.
105. Lehmenkuhler A. Interrelationships between DC potentials, potassium activity,p02 and pC02 in the cerebral cortex of the rat // Origin of fields potentials: Intern. Symposmm, Muenster Germany-Stuttgart-1979-P.49-50.
106. McFarland W.L., Levin S. Electroencephalographic responses to 2500 rads of whole-body gamma-neutron radiation in the monkey Macaca mulatta // Radiat. Res. -1974.-V. 58, N1.- P.60-73.
107. Nandy К. Centrophenoxine: effects on aging mammalian brain. // J. Amer. Geriat. 1978. V. 26. P. 74-81.
108. Nakagawa N. Studies on changes in trace elements of the brain related to aging. // Hokkaido Igaku Zasshi. – 1998. V.73. I. 2. P. 181-199.
110. Neveu P.J., Barneod P., Vitiello S., LeMoal M. Brain modulation of the immune system: association between lymphocyte responsiveness and paw preference in mice // Brain Res. 1988. V. 457. № 2. P. 392.
111. Neveu P.J., Betancur C., Barneod P. et al. Functional brain asymmetry and murine systemic lupus erythematosis // Brain Res. 1989. V. 498. № 1. P. 159.
112. Palm R., Wahlstrom G., Hallmans G. Age related changes in weight and the concentrations of zinc and cooper in the brain of the adult rat // Lab Anim. – 1990. – V.24, Is. 3. – P. 240-5.
113. Pausescu E., Paun C., Teodosiu T. Efecte ale expunerii la radiatii ionizante aspura ultrastructuru si metabolismului creierului de mamifere // Rev. Sanit. Milit. -1974.- N4.-P.325 -334.
114. Rajan K.S. Distribution of metal ions in the subcellular fractions of several rat brain areas // Life Sci. –1976.– V. 18. № 4. –Р.423.
115. Roy D., Pathak D. N., Singh R. Effect of centrophenoxineon the antioxidative enzymes in varios regionsof the aging rat brain. // Exper. Gerontology. –1983.– V. 18, № 3.– Р. 185-187.
116. Sandstead H.H. A brief history of the ifluence of trace elements on brain function // Am. J Clin Nutr. – 1986.– V.43, Is. 2.– P.293-298
117. Somjen G.G, Rosenthal M. Evoked sustained potentials and oxidation of intramitochondrial enzymes in normal and in pathological states // Origin of fields potentials: Intern. Symposium, Muenster, Germany.-Stuttgart.-1979.-P. 164-174.
118. Takeda, A., Akiyama, T., Sawashita, J., Okada, S. Brain uptake of trace metals, zink and manganese, in rats // Brain Res. - V. 640, I. 1-2, P. 341-344.
119. Talan M.I., Engel B.T. Temporal Decreaseof Body Temperture in Middle-aged C57BL/6J MiceY// J. ofGerontol.-1986-V. 41, N 1-P. 8-12.
120. Van Bel F., Shadid M., Dorrepaal C.A, Fontijn J. and all. Effect of allopurinol on postasphyxial free radical formation, cerebral hemodinamics, and electrical brain activity//Pediatrics.-l 998.-N101 .-P. 185-193
121. Wheller K.T., Wierowski S.V., Ritter P. Are inducible components involved in repaire irradiated neuronal and brain tumor DNA? // Int. Radiat. Biol. – 1981. – V. 40. – P.293.