ПРИРОДА ФУНКЦИОНАЛЬНОЙ МОТОРНОЙ АСИММЕТРИИ У ЖИВОТНЫХ
М.Е. Иоффе, Е.В. Плетнева, И.С. Сташкевич
Институт высшей нервной деятельности и нейрофизиологии РАН, Москва
Предпочтение одной из передних конечностей в манипуляционных движениях у животных есть результат взаимодействия исходного предпочтения и обучения. Природа исходного предпочтения не ясна. При селекции мышей по признакам «правшества» и «левшества» обнаружено, что наследуется не знак, а степень моторного предпочтения. Существует, однако, корреляция знака предпочтения с рядом морфологических, функциональных и нейрохимических признаков. Обнаружены популяционные сдвиги знака асимметрии в некоторых линиях мышей. Существуют различия между «правшами» и «левшами» в социальном поведении и в характере обучения и устойчивости к переучиванию у крыс. Линии крыс с разными формами генетической эпилепсии характеризуются преобладанием животных с определенным знаком моторного предпочтения. Эти результаты не позволяют полностью исключить генетические влияния на возникновение исходного предпочтения. Вероятно, среди факторов, определяющих моторное предпочтение, генетические влияния и воздействия среды находятся в тесном взаимодействии.
Проблема функциональной моторной асимметрии у животных составляет часть общей проблемы асимметрии организации и функций мозга, куда входит также морфологическая асимметрия различных структур мозга, химическая асимметрия содержания медиаторов, метаболитов и других веществ, асимметрия межполушарных взаимоотношений и электрической активности мозговых структур и т.д. Функциональная моторная асимметрия включает различные двигательные проявления, как, например, пространственная асимметрия (предпочтение правой или левой стороны при локомоции, плавании, выборе рычага), асимметрия позного тонуса в норме и после унилатеральных повреждений мозга, предпочтительное использование правых или левых конечностей в манипуляционных движениях и т.д. В настоящей работе мы в основном рассмотрим лишь последний вид функциональной моторной асимметрии[1].
Асимметрия при выполнении манипуляционных движений у животных стала систематически исследоваться в 30-е годы ХХ в., когда были получены основные результаты о характере предпочтения конечности у крыс и обезьян и показано, что: 1) в отличие от человека, число “правшей” и “левшей” у животных примерно одинаково ( G.M. Peterson, 1934; L.S. Tsai et al.,1930); 2) индивидуальное предпочтение при выполнении определенной двигательной задачи довольно стойко и не меняется в течение длительного времени ( G.M. Peterson, 1934); 3) в то же время одно и то же животное может менять предпочитаемую конечность при смене двигательной задачи (G.M. Peterson, 1934; J.M. Warren, 1980); 4) степень выраженности предпочтения зависит от сложности и характера выполняемого движения (J.S. Kounin, 1938); 5) предпочтение в значительной степени зависит от обучения и может быть переделано путем специальной тренировки (G.M.Peterson, 1951; K.L. Wentworth, 1942). В дальнейшем эта проблема исследовалась довольно интенсивно (В.Л. Бианки, 1985; R.L. Collins, 1968-1991; V.H. Denenberg, 1981; V.H. Denenberg et al., 1991; R.A.W. Lehman, 1970; D. Martin, W.G. Webster, 1974; J.M. Warren, 1980; и др.). В качестве экспериментальной модели чаще всего использовался предложенный Д.Петерсоном (1934), но нередко называемый тестом Коллинза (Р.Коллинз (1968) применил этот тест при исследовании “рукости” у мышей[2], cм. J.M.Waters, V.H. Denenberg, 1994) тест доставания пищевых шариков из узкой трубки, обычно горизонтальной, но иногда (В.Л. Бианки, 1985) – вертикальной. Исследования были сосредоточены на нескольких аспектах проблемы. Рассмотрим их подробнее.
Предпочтение конечности – результат обучения или индивидуальная особенность?
Одним из основных вопросов, привлекавших внимание исследователей, является вопрос о врожденном или приобретенном характере предпочтения конечности. Существует множество фактов, свидетельствующих, что направление асимметрии – доминирование правой или левой конечности – может определяться особенностями обучения (В.Л. Бианки 1985; D. Martin, W.G. Webster, 1974; E.I. Miklyaeva et al., 1991; G.M. Peterson, 1934; 1951). Обучение, безусловно, играет огромную роль в формировании реального предпочтения конечности. Следует помнить, что предпочтение наиболее выражено в тонких специализированных движениях, которые тоже формируются в процессе обучения, тогда как при выполнении более простых движений те же самые животные, которые проявляют сильную латерализацию в специализированных движениях, нередко являются амбидекстрами (E.I. Miklyaeva et al., 1991; K.L. Wentworth, 1942). Это может быть связано с тем, что мозговое представительство специализированных выученных движений, как правило, унилатерально, на что указывает отсутствие прямого трансфера этих движений при смене активной конечности, показанное ранее как у животных, так и у человека. Простые синергии обычно представлены билатерально.
Исследователи, пытавшиеся выявить исходное предпочтение при минимальном влиянии обучения (И.С. Сташкевич, А.Д. Воробьева, 1997; E.I. Miklyaeva et al., 1991; K.L. Wentworth, 1942), пришли к разным выводам. Согласно K.L.Wentworth (1942), минимальное число проб для выявления предпочтения составляет 10 успешных взятий пищи. Е.И. Микляева с соавт., 1991 на довольно большом материале (168 животных) пришли к выводу, что у нелинейных крыс предпочтение достаточно уверенно выявляется уже после 3 взятий. В то же время И.С.Сташкевич и А.Д.Воробьева (1997) обнаружили, что до 15% крыс линии Wistar, которые могут проявлять выраженное предпочтение в одном или даже нескольких последовательных блоках по 10 взятий, неоднократно меняют его при дальнейшем обучении до 300 взятий. Авторы считают, что именно эти крысы являются настоящими амбидекстрами и для их выявления необходимо длительное обучение. Таким образом, для выявления моторного предпочтения необходимо обучение. В связи с этим еще относительно недавно существовала точка зрения, что предпочтение конечности у животных изначально не существует, а полностью формируется в процессе обучения (С. Спрингер, Г. Дейч, 1983). В самом деле, в процессе формирования специализированного движения доставания пищи из трубки животное, как правило, сначала пытается достать пищу языком, затем совершает быстрые альтернирующие просовывания обеих лап и лишь через некоторое время начинает брать пищу преимущественно одной конечностью. Согласно закону эффекта Торндайка частота движений, приводящих к подкреплению, должна увеличиваться (основной принцип инструментального обучения – “что подкрепляется, то закрепляется”). Иначе говоря, первое успешное движение правой или левой лапой должно привести к тому, что в дальнейшем животное будет использовать в данном тесте преимущественно эту конечность. Такой подход полностью объяснял формирование предпочтения конечности, не требуя привлечения генетических и других возможных механизмов.
Для проверки предположения, что моторное предпочтение целиком является результатом обучения, были проведены специальные эксперименты (E.I. Miklyaeva et al., 1991). После минимального первичного обучения и выявления предпочитаемой конечности (3 успешных доставания пищи из горизонтальной трубки) у нелинейных крыс электрошоком вызывали ретроградную амнезию. О степени амнезии судили, сравнивая время повторного обучения с временем первичного обучения. У интактных животных и крыс с неполной амнезией время повторного обучения укорачивается. Отсутствие такого укорочения говорит в пользу полной амнезии. Через 2-4 недели после полной амнезии (25 крыс) при тестировании до 3 или до 10 взятий пищи предпочтение сохранилось у 22 животных (88%), 2 крысы (8%, амбидекстры при первичном обучении) приобрели предпочтение, 1 (1,7%) потеряла предпочтение (стала амбидекстром) и лишь 1 крыса (1,7%) сменила предпочитаемую конечность. Эти результаты явно свидетельствовали в пользу точки зрения, что кроме обучения существует исходное предпочтение, которое выявляется при обучении. Правда, некоторая слабость этих экспериментов состоит в том, что степень амнезии определялась по косвенному признаку (время повторного обучения).
Таким образом, реальное предпочтение конечности в манипуляционных движениях, очевидно, определяется двумя факторами – исходным предпочтением и обучением. Было показано, что степень исходного предпочтения в популяции нелинейных крыс варьирует у разных животных. Так, при обучении животных доставать пищу только одной (например, левой) лапой (у отверстия трубки ставили вертикальную перегородку, препятствующую просовываниям другой конечности) без предварительного определения их исходного предпочтения, разные животные вели себя по-разному: одни крысы обучались легко (предположительно, первичные левши и амбидекстры), другие – с трудом (предположительно, первичные правши, у которых в процессе обучения происходила переделка исходного предпочтения), а некоторые животные не могли обучиться и даже ложились на спину, чтобы иметь возможность взять пищу правой лапой, исходное предпочтение которой было у них, видимо, крайне высоко (E.I. Miklyaeva et al., 1991).
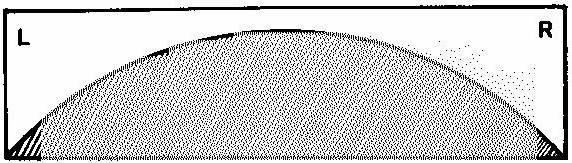
Было высказано предположение, что вся популяция животных представляет собой континуум, в котором на полюсах находятся животные с очень сильным исходным предпочтением той или другой конечности, а промежуток заполнен особями с постепенным снижением предпочтения одной конечности и (через группу амбидекстров) переходом к постепенно увеличивающемуся предпочтению другой лапы. При этом, чем меньше исходное предпочтение у какой-либо особи, тем выше роль обучения в формировании реального предпочтения (рис.1) (E.I. Miklyaeva et al., 1991).
Рис.1. Схема соотношения исходного предпочтения (светлая часть) и обучения (заштрихованная) в реальном предпочтении конечности у животных (Miklyaeva et. al., 1991).
Исходное предпочтение – ригидное врожденное свойство или результат пластических изменений в онтогенезе?
В связи с этим встает вопрос о природе так называемого исходного предпочтения. В.Л.Бианки (1985) отмечал, что у домовой мыши обнаруживается не только индивидуальная, но и популяционная асимметрия, а именно преобладание использования правой конечности в манипуляционных движениях у большинства животных. У кошек в разных манипуляционных тестах наблюдалось преобладание правшей (Г.М. Глумов, 1984) или левшей (M. Fabre-Thorpe et al., 1993). Все это могло бы говорить в пользу врожденного характера моторного предпочтения. Попытки прямого решения вопроса о генетическом характере предпочтения путем скрещивания крыс или мышей с одинаковым предпочтением (В.Л. Бианки, 1985; R.L. Collins, 1985-1991; G.M. Peterson, 1934) показали, что знак моторного предпочтения (доминирование правой или левой конечности) не наследуется, однако наследуется степень его выраженности, т.е. частота использования предпочитаемой конечности в манипуляционных движениях. Последнее наблюдение было неоднократно подтверждено (F.G. Biddle, B.A. Eales, 1999; P. Signore et al., 1991). Получены и широко исследуются сильно и слабо латерализованные по моторному предпочтению линии мышей (F.G. Biddle, B.A. Eales, 1999; R.L. Collins, 1985; D. Martin, W.G. Webster, 1974). Дискутируется вопрос, контролируется ли степень предпочтения одним или несколькими генами (F.G. Biddle et al., 1993; F.G. Biddle, B.A. Eales, 1996) или она связана с разной степенью гетерозиготности (мыши с большей гетерозиготностью обнаруживают большую степень предпочтения конечности) (I.C. McManus, 1992).
Таким образом, до настоящего времени нет прямых доказательств наследственной передачи правшества и левшества у животных. В то же время показано, что и степень латерализации в одной и той же линии животных зависит не только от генетических факторов, но и от процедуры тестирования. Так, после редких, но длительных тестирований (по 2 взятия в день в течение 25 дней) частота использования предпочитаемой лапы была существенно больше, чем после более интенсивных, но кратковременных (по 25 взятий в день в течение 2 дней) (M.B. Bulman-Fleming et al., 1997).
Логично предположить, что, поскольку знак предпочтения не наследуется, видимо, это не врожденное свойство особи, а результат раннего онтогенетического опыта (V.H. Denenberg, 1981, 2000), прежде всего воспитания в асимметричной среде (В.Л. Бианки, 1985; R.L. Collins, 1985; R.A.W. Lehman, 1970). Действительно, обычно трудно полностью исключить асимметричную среду. Даже если в одной клетке воспитывается несколько особей, проявляющих разное предпочтение, всегда остается некоторая возможность того, что животные, занимающие в иерархии подчиненное положение, оттесняются доминантами в углы и к стенкам клетки, что приводит к неравному использованию правых и левых конечностей и в конце концов к формированию разного предпочтения у разных животных.
Асимметричная среда может возникать и в антенатальном онтогенезе, например разное положение плодов в матке могло бы создавать разные условия для развития и использования конечностей. Можно найти и ряд других факторов среды, которые могли бы быть причиной функциональной моторной асимметрии. Показано, в частности (V.H. Denenberg et al., 1991), что аутоиммунные процессы в матке матери усиливают степень, но не определяют знак моторного предпочтения у самок (но не у самцов) мышей, развивавшихся до рождения в соответствующей среде. Последний пример показывает, что даже симметричная среда может влиять на развитие асимметрии мозга и/или предпочтения одной из передних конечностей. Другим примером является латерализация полушарий мозга у крысят вследствие раннего хэндлинга (взятия на руки), что приводило к мощным перестройкам в организме детеныша и, в частности, к развитию межполушарного торможения. Специальными экспериментами было показано, что у взрослых крыс, подвергавшихся хэндлингу в раннем возрасте, в тесте открытого поля правое полушарие тормозит левое, а в тесте на направление первой побежки – наоборот (V.H. Denenberg, 2000). Измененное вследствие раннего опыта поведение самок, в частности материнское поведение, могло влиять и на последующие поколения. Таким образом, влияния среды очень сложны и нередко неожиданны, что крайне затрудняет деление фенотипических признаков на врожденные и приобретенные.
Пытаясь все же как-то проанализировать вопрос о генезе «исходного» моторного предпочтения, рассмотрим ряд исследований, результаты которых свидетельствуют о связи знака предпочтения (правшество – левшество) с рядом разнообразных функциональных и морфологических особенностей организма, что как будто говорит против предположения о возникновении предпочтения в онтогенезе только в результате влияния среды.
Корреляция «рукости» с морфологическими факторами. Рядом работ установлено, что некоторые структурные изменения мозга у животных разных видов коррелируют с предпочтением определенной конечности в манипуляционных движениях. Показано, например, наличие корреляции между моторной асимметрией и различиями веса полушарий мозга у кошек (U. Tan, N. Kutlu, 1993). Обнаружено, что направление моторного предпочтения отрицательно коррелирует с генетически обусловленной морфологической асимметрией сенсорной системы вибрисс у мышей (P. Barneod, H. Van der Loos, 1993). Авторы задаются вопросом, связаны ли изменения систем мозга, обусловливающие асимметрию чувствительной системы вибрисс, с изменениями, обусловливающими моторное предпочтение, или существуют два ассоциированных набора генов, один из которых обусловливает «вибриссность», а другой «рукость». Возможно, правда, и еще одно альтернативное объяснение, связанное с тем, что одностороннее снижение вибриссной чувствительности может привести к компенсаторному повышению тактильной чувствительности с этой стороны и, как следствие, к преимущественному использованию соответствующей конечности в манипуляционных движениях.
В другой работе исследована связь асимметрии интра- и инфрапирамидных проекций мшистых волокон гиппокампа с направлением и знаком моторного предпочтения у мышей разных линий (H.P. Lipp et al., 1996). Прежде всего было установлено, что проекции мшистых волокон гиппокампа в общей популяции у мышей сильно латерализованных линий на 70% мощнее, чем у животных слабо латерализованных линий (и у мышей с дисплазией мозолистого тела), и показана положительная корреляция между индивидуальным развитием проекций мшистых волокон и степенью моторного предпочтения. Особенно интересно, что асимметрия пучков мшистых волокон достоверно коррелировала и с направлением моторной асимметрии, т.е. с правшеством или левшеством. Авторы приходят к заключению, что степень выраженности и асимметрия площади проекций пучков мшистых волокон гиппокампа является одним из многих факторов, влияющих на направление и силу моторного предпочтения. Пытаясь объяснить полученные результаты, авторы предполагают, что животные с более мощными пучками мшистых волокон более устойчивы к интерферирующим влияниям и поэтому чаще употребляют определенную лапу (высокая степень асимметрии), тогда как право-левая асимметрия гиппокампальных пучков может влиять на начальный выбор лапы и/или подкреплять (?) его.
У мышей линии BALB/cCF обнаружена связь между степенью развития мозолистого тела и направлением моторного предпочтения (S.L. Schmidt et al., 1991). В группе животных с полной или частичной агенезией мозолистого тела процент левшей (78%) был достоверно выше, чем при случайном распределении, тогда как у здоровых животных распределение правшей и левшей не отличалось от случайного. В то же время различий между группами по степени моторного предпочтения не обнаружено.
Таким образом, ряд структурных изменений мозга коррелирует с направлением функциональной моторной асимметрии.
Иммунный и гормональный статус и моторное предпочтение. Известно, что морфологические изменения могут быть связаны с гормональными сдвигами, например, при акромегалии. Возможно поэтому, что описанные выше структурные изменения и коррелирующее с ними направление исходного моторного предпочтения (правшество или левшество) могут в какой-то части, например в случае с агенезией мозолистого тела, быть результатом нарушений гормонального статуса. Действительно, в ряде исследований показана корреляция гормональных сдвигов со знаком моторного предпочтения. Обнаружены разные изменения содержания гипофизарного гормона пролактина в ответ на стресс у животных с разным моторным предпочтением, причем эти изменения были различны в разных генетических линиях (N.S. Waters et al., 1996). В генетически гетерогенной группе, полученной от скрещивания мышей аутоиммунной линии NZB и гетерозиготной по окраске линии SM, у самок мышей черной окраски предпочтение левой лапы в тесте Коллинза коррелировало с достоверным увеличением содержания пролактина, а у правшей нет. Среди самок окраски агути, напротив, содержание пролактина было достоверно снижено у левшей, а у правшей нет. В генетически гомогенной группе C3H/HeJ только самки-правши реагировали на стресс существенным повышением содержания пролактина. В то же время самцы ни в одной из групп не обнаруживали корреляции моторного предпочтения с изменением содержания пролактина в ответ на стресс. Подобные же результаты (корреляция с рукостью только у самок) были получены при исследовании содержания АКТГ (C. Betancur et al., 1992).
У мышей обнаружена связь между интенсивностью иммунных реакций и знаком моторного предпочтения (C. Betancur et al., 1991; E. Fride et al., 1990; P.J. Neveu, 1991; P.J. Neveu et al., 1988, 1989). Результаты различались у животных разных линий. У леворуких самок мышей линии C3H/HeJ пролиферация лимфоцитов более интенсивна, чем у праворуких (P.J. Neveu et al., 1988), а у леворуких самцов линии C3H/OuJIco активность лимфоцитов-киллеров выше, чем у праворуких (C. Betancur et al., 1991). Активность антител у леворуких самок в аутоиммунной линии NZB выше, чем у праворуких (P.J. Neveu et al., 1989). Хотя некоторые авторы (P.J. Neveu, 1991) подчеркивают более активный иммунный статус у левшей, другие (E. Fride et al., 1990) обращают внимание на различия иммунных реакций в разных линиях мышей. Так, в линиях C3H/HeJ и 129/J леворукие мыши демонстрировали более сильную иммунную реакцию, а в линиях C3H/HeNCr MTV- и BALB/cJ – наоборот. В линиях C3H/HeNCr MTV+ и в гетерогенной контрольной популяции различий между правшами и левшами по интенсивности иммунных реакций не обнаружено (E. Fride et al., 1990). Таким образом, связь между иммунными реакциями и моторным предпочтением зависит от линии животных, что позволяет думать о генетических влияниях на ее природу.
Различия между линиями животных по направлению моторного предпочтения. Существуют и прямые данные о преобладании различного предпочтения в разных линиях мышей. При исследовании предпочтения в большой популяции мышей (n=693) 29 инбредных линий (N.S. Waters, V.H. Denenberg, 1994) были обнаружены различия между линиями по направлению моторной асимметрии в одном из двух применявшихся тестов. В другой работе (F.G. Biddle et al., 1993) при исследовании 12 инбредных линий мышей в некоторых линиях был обнаружен достоверный сдвиг в сторону предпочтения правой или левой лапы, не зависимый от степени латерализации, что, по мнению авторов, говорит о том, что направление предпочтения может быть отдельным генетическим признаком в использовавшихся экспериментальных моделях. Тем не менее селектировать линии правшей и левшей не удается.
Рассмотрим теперь некоторые особенности поведения, связанные с моторным предпочтением.
Моторное предпочтение и социальное поведение.
Коротко остановимся на социальном поведении, которое, с одной стороны, представляет собой развитие генетически предетерминированной программы, а с другой – сильно модулируется условиями среды. Показано (Е. И. Варлинская c соавт., 1993), что нарушения социального поведения, возникающие после изоляции беспородных взрослых крыс-самцов, различны, в зависимости от моторного предпочтения. У “абсолютных правшей” (10 взятий пищи правой лапой в 10 пробах) и амбидекстров изоляция увеличивала уровень агрессии, а у “абсолютных левшей” – внутривидовых контактов. При этом у всех животных нарушается видотипическая структура агрессивного поведения. Авторы связывают полученные данные с разной ролью правого и левого полушарий в регуляции агрессивного поведения и его структуры. При этом априори предполагается, что у левшей активнее правое полушарие, а у правшей левое и что моторная и эмоциональная доминантность полушарий совпадают.
Обучение у животных с разным моторным предпочтением.
Рассмотрим теперь, как исходное моторное предпочтение влияет на обучение манипуляционным движениям. Частично этот вопрос уже затрагивался выше. Полученные недавно данные говорят о том, что животные с разным предпочтением обучаются по-разному. Проблема таких исследований состоит в том, что нельзя определить исходное предпочтение без обучения, которое само модулирует предпочтение. Однако можно подойти к проблеме с другой стороны – ретроспективно проанализировать начало и дальнейший ход обучения у животных с разным финальным предпочтением, сложившимся в процессе обучения. Оказалось (И.С. Сташкевич, М.А. Куликов, 2000), что распределения коэффициента асимметрии,
П - Л
вычисленного из 10 последовательных проб по формуле Кас= ---------,
П + Л
где П – число взятий пищи правой лапой, а Л – число взятий левой лапой, различаются на ранних стадиях обучения у крыс линии Wistar (n=61), которые в конце обучения становятся левшами и правшами. Как видно из рис.2, в первых 10 пробах будущие правши и левши, на первый взгляд, распределяются довольно равномерно. Часть из них не проявляет выраженного предпочтения (коэффициент асимметрии колеблется в пределах от –0,4 до 0,4), а число абсолютных правшей и абсолютных левшей (коэффициент асимметрии равен ±1) примерно одинаково.
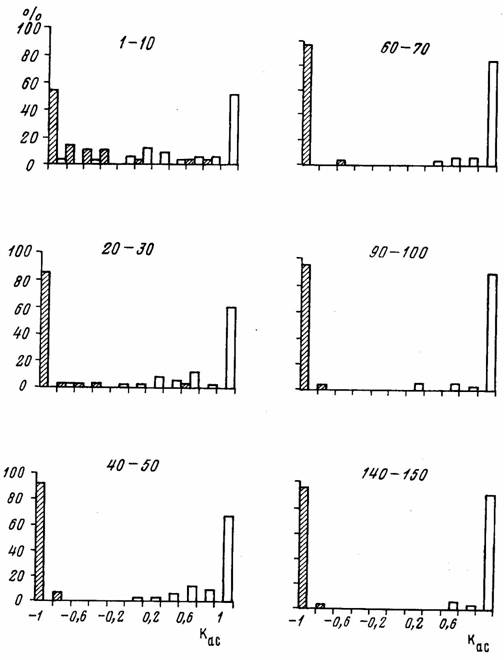
Рис.2. Распределение величин коэффициента асимметрии в процессе обучения у крыс, ставших после обучения «правшами» (светлые столбики) и «левшами» (заштрихованные).
По оси абсцисс - величины коэффициента асимметрии, по оси ординат - процент животных (И.С.Сташкевич, М.А.Куликов, 2000). Цифры над столбиками – последовательные пробы.
Однако более детальный анализ показывает, что распределения будущих правшей и будущих левшей различаются (p<0.003). Распределение будущих правшей более растянуто и бимодально. Видно также, что несколько животных из обеих групп имеют противоположное предпочтение на ранней стадии обучения. При дальнейшем обучении группа левшей формируется быстрее правшей. Различия между распределениями достоверны (p<0.001) после 30, 50 и 70 проб. После 70 проб практически все левши (96%) достигают максимального уровня латерализации, который не меняется при дальнейшей тренировке, тогда как уровень полной латерализации правшей все еще ниже (85%) и только после 100 проб 91% правшей достигают максимальной латерализации. Таким образом, при длительном обучении левши обучаются быстрее правшей.
Взаимодействию исходного предпочтения и обучения посвящен также ряд работ, касающихся переделки предпочтения путем ограничения возможности выполнять движение предпочитаемой конечностью (D. Martin, W.G. Webster, 1974, E.I. Miklyaeva et al., 1991; E.I. Miklyaeva, J. Bures, 1991; R. Milisen, 1937; G.M. Peterson, 1951; K.L. Wentworth, 1942). Результаты этих работ свидетельствуют, что для смены предпочтения требуется новое обучение (forced practice – термин, введенный Д.Петерсоном (1951)). При этом переделка была более или менее трудной в зависимости от силы исходного предпочтения (E.I. Miklyaeva et al., 1991). Было показано (Ю.В. Васильева с соавт., 1995), что беспородные крысы-правши после кратковременного обучения (10 успешных проб) переучиваются легче, чем левши по показателям времени, необходимого для совершения первого успешного взятия пищи, числа подкреплений за 1 мин (рис.3) и числа попыток использовать исходно предпочитаемую лапу.
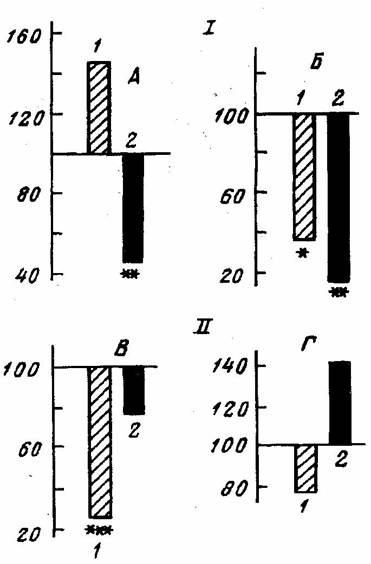
Рис.3. Сравнение длительности одного из периодов обучения (I) и скорости взятий подкрепления (II) при переобучении с показателями при обучении, принятыми за 100%. А, В – левши (1) и правши (2); Б, Г – амбидекстры, обучившиеся использовать только правую (1) и левую (2) лапу. * - p<0.05, ** - p<0.01, *** - p<0.001 (Ю.В.Васильева и др., 1995).
В то же время анализ этого феномена у крыс линии Wistar в условиях интенсивного обучения (300 проб) и длительной переделки предпочтения (300 проб) (И.С. Сташкевич с соавт., 2001) показал, что после возврата к свободному выбору конечности животные делятся на несколько групп: 34% крыс сохранили навязанное предпочтение, 23% стали амбидекстрами, а 43% вернулись к первоначальному предпочтению. В каждой из групп было примерно одинаковое число правшей и левшей, однако первоначальные левши проявляли более выраженную тенденцию использовать первично предпочитаемую лапу, чем правши, что наиболее четко видно в группе животных, ставших амбидекстрами (рис.4).
Рис.4. Динамика использования правой и левой конечности у животных – исходных правшей и левшей, после переобучения и возвращения к свободному выбору конечности. I – первичное обучение, II – переобучение, III – свободный выбор после переобучения. По оси абсцисс – блоки по 50 проб, по оси ординат - процент животных. * - p<0.05, ** - p<0.01, *** - p<0.001 (И.С.Сташкевич и др., 2001).
Таким образом, крысы обучаются двигательному навыку с разной скоростью и проявляют индивидуальные различия устойчивости к переделке, что, очевидно, связано с разной степенью исходного предпочтения. При этом, однако, обнаруживаются различия правого и левого полушарий в организации исходного предпочтения, проявляющиеся при обучении и переделке. Нельзя исключить, что этот популяционный признак может иметь генетическую основу. Одно из возможных объяснений природы этих межполушарных различий таково. В процессе обучения почти всегда имеется стадия бимануальных альтернирующих движений, после которой животное начинает преимущественно использовать одну лапу. Очевидно, это связано с формированием торможения движений другой конечности (возможно, транскаллозальным (S. Levitan, J.A. Reggia, 2000)). Можно предполагать, что такое межполушарное торможение сильнее выражено при исходном моторном доминировании правого полушария, т.е. у потенциальных левшей, которые быстрее формируют реакцию одной конечности, труднее переделывают ее и проявляют более выраженную тенденцию к возврату к первично предпочитаемой конечности после длительной переделки. Следует, однако, помнить, что и направленное межполушарное торможение может быть результатом влияний среды, как в приводимых выше экспериментах с ранним хэндлингом.
Показано, что переделка предпочтения (обучение в камере со смещенной в одну сторону трубкой) происходит по-разному у мышей разных генетических линий (S. Betancur et al., 1992). Мыши сильно латерализованной линии C57BL/GJ достаточно быстро обучались использовать определенную конечность, тогда как животные слабо латерализованной линии CDS/Lay не переучивались, оставаясь стойкими амбидекстрами. В то же время из двух других слабо латерализованных линий SWV и NOD/Lt первая оказалась переучиваемой, а вторая – нет. Авторы заключают, что вероятность “правшества” или “левшества” зависит как от генетических факторов, так и от характера теста.
Обучение другим навыкам происходит по-разному у животных-правшей и левшей. Так (V.H. Denenberg et al., 1991), среди мышей аутоиммунной линии NZB левши (как самцы, так и самки) быстрее правшей обучались находить безопасную платформу при плавании. Та же картина наблюдается у мышей линии BXSB, но только у самцов. Однако результаты дискриминативного обучения у самцов линии BXSB были противоположными – правши обучались быстрее левшей.
Таким образом, животные с разным моторным предпочтением и с разными генетическими особенностями по-разному обучаются, по-разному переучиваются и обнаруживают разную устойчивость исходного предпочтения к переделке.
Экспериментальная патология и моторное предпочтение.
Большой интерес представляют исследования моторного предпочтения у животных с экспериментальной патологией. В течение ряда лет группа В.Дененберга исследует моторную асимметрию и обучение у мышей аутоиммунных линий (V.H. Denenberg, 2000; V.H. Denenberg et al., 1991a,b, 1992). Целью этих исследований является выяснение соотносительной роли генетических факторов и раннего опыта, в частности, в формировании моторного предпочтения. Показано (V.H. Denenberg et al., 1991a), что после пересадки оплодотворенной яйцеклетки здоровой мыши (линия DBA) в матку мыши аутоиммунной линии BXSB потомство (самки) обнаруживали повышение титра антител, увеличение степени моторной асимметрии и нарушения выработки реакций избегания и избавления. В то же время пересадка яйцеклетки аутоиммунной мыши в матку неаутоиммунной гибридной мыши приводила к обратным результатам. Авторы заключают, что описанные нарушения носят эпигенетический характер и подчеркивают роль среды, в которой развивается эмбрион и плод, в возникновении аутоиммунных механизмов. С другой стороны, возникновение корковых эктопий у аутоиммунных мышей имеет генетическую природу.
Среди моделей экспериментальной патологии представляют интерес различные формы эпилепсии. В то же время до последнего времени не исследовалась моторная асимметрия у животных-эпилептиков. Недавно (Е.В. Плетнева, 1999; Е.В. Плетнева, М.Е. Иоффе, 2000) были обнаружены разнонаправленные популяционные сдвиги моторного предпочтения у крыс с разными формами эпилепсии. Оказалось, что среди крыс линии WagRij, характеризующихся наличием absence-эпилепсии с пик-волновыми комплексами в корковой ЭЭГ, достоверно преобладают левши, тогда как в группе крыс WagRij со смешанной формой эпилепсии, у которых наблюдаются как absence-эпилепсия, так и аудиогенные судороги, доминируют правши. Преобладание правшей было обнаружено и в других линиях крыс с аудиогенной судорожной эпилепсией – в эпилептической популяции крыс Wistar (Е.В. Плетнева, М.Е. Иоффе, 2000) и у крыс линии КМ (Е.В. Плетнева, М.Г. Плескачева, 2001) (таблица). Оказалось при этом, что степень сдвига вправо у крыс линии Wistar прямо коррелирует с интенсивностью судорожного припадка (Е.В. Плетнева, М.Г. Иоффе, 2000). В потомстве крыс с аудиогенными судорогами процент животных с высокой интенсивностью припадка возрастает, соответственно возрастает и процент правшей. Эти результаты кажутся очень интересными в двух аспектах. Прежде всего, в разных генетических линиях крыс имеют место разнонаправленные сдвиги моторного предпочтения. Этот результат, как и ряд представленных выше данных, говорит в пользу наличия генетического компонента в моторном предпочтении. Этот компонент у ряда животных ассоциирован с некоторыми другими генетически детерминированными признаками, как например, генетическая эпилепсия. Поскольку генетическая основа высокой аудиогенной чувствительности у крыс в значительной мере исследована, в частности, показана полигенная структура этого признака у крыс КМ (Л.Г. Романова с соавт., 1976), это дает некоторую надежду на прогресс в исследованиях генетической структуры моторного предпочтения. Второй аспект связан с физиологическим анализом разных форм эпилепсии, что может помочь понять природу моторной асимметрии. Известно, например, что пик-волновые комплексы при absence-эпилепсии генерируются таламическими структурами (см. M. Steriade, M. Deschenes, 1984 и др.), а аудиогенную эпилепсию связывают с патологией задних бугров четверохолмия (J. Tsutsui et al., 1992; и др.). Кроме того, возможно, разные формы эпилепсии имеют разную нейрохимическую основу. В этом случае, зная ее, можно попытаться подойти к анализу разной нейрохимической природы разнонаправленного моторного предпочтения (конечно, если принять соответствующую гипотезу).
К сожалению, хотя нейрохимические механизмы эпилепсии активно изучаются, различия между формами эпилепсии не до конца ясны. Считается, что в основе эпилептогенеза лежит дисбаланс тормозных (ГАМК) и возбуждающих (глютамат) медиаторов. Известны антисудорожные эффекты потенциации прямых ГАМКергических (A. Depaulis et al., 1994) и блокады непрямых глютаматергических (C. Deransart et al., 1996) проекций стриатума в ретикулярную часть черной субстанции. Показана роль передачи дофамина в n.accumbens в возникновении absence-эпилепсии (C. Deransart et al., 2000). Введение агонистов дофаминовых Д1 и Д2 рецепторов уменьшало, а введение антагонистов увеличивало выраженность absence-эпилепсии. Эффект, по-видимому, опосредован таламическими проекциями базальных ганглиев. Обнаружено увеличение выделения ГАМК и глютамата в вентробазальном ядре таламуса при генерации absence-эпилепсии (P.K. Banerjee, O.K. Sneed, 1995). Таким образом, пока можно говорить лишь о разных структурах, преимущественно связанных с возникновением разных форм эпилепсии.
Нейрохимическая асимметрия и моторное предпочтение.
Приведенные выше данные наводят на мысль, что моторное предпочтение имеет химическую природу и может быть связано с химической асимметрией мозговых структур. Известна химическая асимметрия нигро-стриарной системы (S. Cabib et al., 1885; S.D. Glick et al., 1977; D. M. Nielsen et al., 1997).. Показано асимметричное распределение М-ацетилхолиновых рецепторов в коре крысы (M.F. Pediconi et al., 1993). Выявлена межполушарная асимметрия активности некоторых ферментов - аденилатциклазы в коре и гиппокампе, а 5΄-нуклеотидазы – в амигдале (И. А. Журавин с соавт., 1995). Показано, что внутриутробная гипоксия приводит к более сильным нарушениям функционального состояния клеточных мембран (по уровню активности ряда ферментов) правого полушария эмбрионов по сравнению с левым (Н.Н. Наливаева с соавт., 1998). Обнаружены асимметрично действующие пептиды (G.Ya. Bakalkin, 1989), например вазопрессин (Г.А. Вартанян, Б.И. Клементьев, 1991).
Особый интерес вызывают исследования химической асимметрии в связи с моторным предпочтением. Обнаружено большее содержание дофамина в n.accumbens, ипсилатеральном предпочитаемой конечности (S. Cabib et al., 1995). Показано (D.M. Nielsen et al., 1997), что интенсивность спонтанных вращательных движений и степень предпочтения одной конечности у сильно латерализованной линии мышей коррелируют с асимметрией дофамина и его метаболита дигидроксифенилуксусной кислоты соответственно в медиальной префронтальной коре. Хотя знак предпочтения конечности и направление вращательных движений не коррелируют между собой, мыши с противоположным направлением этих видов моторной асимметрии обнаруживают различия метаболизма дофамина в n.accumbens и в стриатуме. Обнаружены различные изменения активности ацетилхолинэстеразы, 5΄-нуклеотидазы и аденилатциклазы в сенсомоторной коре крыс на 2-й и 45-й день после кратковременного обучения доставать пищу из трубки (10 проб) (Н.Н. Наливаева с соавт.,1996). По активности всех исследуемых ферментов в обоих полушариях на 2-й день после обучения амбидекстры отличались от животных с выраженным предпочтением, а правши отличались от левшей. Через 45 дней обнаружены различия активности 5΄-нуклеотидазы и аденилатциклазы между амбидекстрами и животными с выраженным предпочтением. Асимметрия активности показана у амбидекстров (ацетилхолинэстераза, 45-й день), у правшей (5΄-нуклеотидаза, 45-й день) и у левшей (аденилатциклаза, 2-й день).
Таким образом, моторное предпочтение действительно коррелирует с химической асимметрией мозга. Однако вопрос осложняется тем, что предпочтение выявляется только при обучении, а обучение, так же как и преимущественное использование одной конечности, даже при 10 пробах сами по себе могут сопровождаться асимметричными химическими изменениями в структурах мозга. Пока не ясно, можно ли и каким образом отдифференцировать химическую асимметрию, возникающую при обучении и тренировке, от асимметрии, связанной с исходным предпочтением. А именно природа исходного предпочтения представляет основной интерес.
ЗАКЛЮЧЕНИЕ: В ПОИСКАХ РЕШЕНИЯ
В настоящей статье мы фиксировали внимание на двух существенных вопросах, интересующих исследователей обсуждаемой проблемы: генетический или приобретенный характер моторного предпочтения и его возможная химическая природа. По поводу первого следует сказать, что в последнее время альтернатива «врожденное – приобретенное» все больше заменяется принципом взаимодействия и взаимной модификации врожденных и приобретенных форм поведения (З.А. Зорина c соавт., 1999; Д. Мак-Фарленд, 1988; V.H. Denenberg, 2000). Хотя структура генов очень консервативна, их функция может изменяться в процессе эволюции. Показателен пример, приведенный В. Дененбергом (V.H. Denenberg, 2000): мышь и человек имеют примерно то же число генов, причем 94% из них идентичны у обоих видов. Учитывая огромную разницу в анатомии, физиологии и поведении, приходится допустить, что идентичные гены могут обеспечивать разные функции. Существенную роль в развитии генетически детерминированных признаков играет как взаимодействие генов между собой, так и их взаимодействие со средой. В связи с этим классическое противопоставление “nature-nurture” ныне считается устаревшим. Среда может существенно изменить проявление генетически детерминированной поведенческой программы. Согласно выражению В. Дененберга (2000), «эволюция предполагает, а онтогенез располагает». С другой стороны, адаптивные пластические изменения, возникшие в процессе жизни индивидуума, могут закрепляться в процессе эволюции.
Выше приводились примеры очень сложных воздействий среды, влияющих на асимметричное поведение, таких как действие внутриматочной среды на плод, ранний хэндлинг детеныша или даже его предков. С другой стороны, приведенные факты множественных корреляций знака моторного предпочтения с морфологическими, физиологическими, биохимическими, генетическими особенностями организма и с некоторыми патологическими процессами не позволяют полностью исключить генетический механизм. Вероятно, среди факторов, определяющих моторное предпочтение, генетические влияния и влияния среды находятся в тесном взаимодействии. Это не исключает интереса к выделению каждого из этих компонентов, однако крайне затрудняет исследование данного вопроса. В поисках путей решения представляется интересным, в частности, дальнейший анализ (поведенческий, физиологический, генетический и биохимический) связи между разными формами генетической эпилепсии и моторным предпочтением. Если удастся найти четкие нейрохимические различия между формами эпилепсии, это, среди прочих важных последствий, может дать ключ к разгадке природы моторного предпочтения.
Среди других подходов может оказаться интересным анализ связи моторного предпочтения и других видов моторной асимметрии, химическая природа которых в определенной степени изучена. Имеется в виду асимметрия позы, вызванная асимметричными травмами мозга (A.M. Di Giorgio, 1929). Известно, что эта асимметрия индуцируется химическими факторами, одним из которых является аргинин-вазопрессин, действующий в очень низких дозах (Г.А. Вартанян, Б.И. Клементьев, 1991). Интраамниотическое введение вазопрессина приводит к стойкой асимметрии позы хвоста у потомства крыс (Б.И. Клементьев с соавт., 1995). Анализ возможной роли вазопрессина и других асимметрично действующих пептидов (G.Ya. Bakalkin, 1989) в организации моторного предпочтения кажется весьма интересным.
Существуют и другие актуальные вопросы, касающиеся организации моторного предпочтения, которые остались за рамками настоящей статьи. К ним относится, в частности, структурная организация моторного предпочтения - роль моторной коры, стриатума и других мозговых структур. Известно, что экстирпация моторной коры контралатерально предпочитаемой лапе приводит к смене предпочтения (G.M. Peterson, 1934, 1951), однако после двусторонней экстирпации исходное предпочтение сохраняется. Вероятно, моторная кора связана не с организацией, а с реализацией моторного предпочтения. Вопрос, существует ли структура, ответственная за организацию предпочтения, остается открытым. В тесной связи с этим находится вопрос о соотношении организации и реализации предпочтения (preference and performance). До сих пор не вполне ясно, насколько более совершенно выполнение манипуляционных движений предпочитаемой конечностью, есть ли разница в выполнении движений правой и левой лапой и т.д. Крайне интересен вопрос об асимметрии компенсации после асимметричных повреждений мозга. Показана разница в сроках и степени восстановления латерализованного навыка после право- и левосторонних повреждений моторной коры и стриатума. Механизмы этих различий еще не исследованы. Не ясна еще связь механизмов предпочтения конечности и других латерализованных форм поведения, а также межполушарной асимметрии, выявляемой электрофизиологически. Согласно одной точке зрения уже у крыс и цыплят левое полушарие больше связано с переработкой информации, а правое – с аффективным поведением (V.H. Denenberg, 2000). Не ясно, влияют ли эти межполушарные различия на моторное предпочтение. Интересен вопрос об эволюции моторного предпочтения. Существует обширная литература, касающаяся «рукости» у обезьян. Однако обзор этих аспектов исследования моторного предпочтения должен быть предметом специальной (и, вероятно, не одной) статьи.
Возвращаясь к основному вопросу, обсуждавшемуся в настоящей статье, следует сказать, что представленные в литературе результаты не позволяют полностью исключить генетические влияния на возникновение исходного предпочтения. Вероятно, среди факторов, определяющих моторное предпочтение, генетические влияния и влияния среды находятся в тесном взаимодействии.
Работа выполнена при поддержке Российского фонда фундаментальных исследований (гранты № 02-04-48410 и 01-04-48381).
Бианки В.Л. Асимметрия мозга животных. Л.: Наука, 1985, 295с.
Вартанян Г.А., Клементьев Б.И. Химическая симметрия и асимметрия мозга. Л.: Наука, 1991. 150 C.
Васильева Ю.В., Варлинская Е.И., Петров Е.С. Исследование переделки двигательного навыка у крыс с различным моторным предпочтением // Журн. высш. нервн. деят. 1995. Т.45. № 5. С.1029
Глумов Г.М. Асимметрия двигательной функции передних конечностей у кошек // Физиол. журн. им. И.М.Сеченова. 1984. Т. 70. № 9. С. 1332
Журавин И.А., Наливаева Н.Н., Плеснева С.А. и др. Активность аденилатциклазы и 5΄- нуклеотидазы в сенсомоторных и лимбических структурах мозга крыс после обучения манипуляторному навыку // Росс. физиол. журн. им. И.М.Сеченова. 1995.Т 81. № 1. С. 40.
Зорина З.А., Полетаева И.И., Резникова Ж.И. Основы этологии и генетики поведения. М.: Изд-во МГУ. 1999. 383 C.
Клементьев Б.И., Рыжова Л.Ю., Игнатьева Т.В. и др. Популяционный профиль асимметрии мозга крыс после внутриамниотического введения вазопрессина // Росс. физиол. журн. им. И.М.Сеченова. 1995. Т.81. № 8. С. 104.
Мак-Фарленд Д. Поведение животных. М: Мир, 1988. 519 C.
Наливаева Н.Н., Плеснева С.А., Чекулаева У.Б. и др. Некоторые биохимические особенности сенсомоторной коры крыс-правшей, левшей и амбидекстров // Журн. эволюц. физиологии и биохимии. 1996. Т.32. № 1. С. 75.
Наливаева Н.Н., Клементьев Б.И., Плеснева С.А. и др. Влияние гипоксии на состояние клеточных мембран правого и левого полушарий мозга эмбрионов крыс // Журн. эволюц. физиологии и биохимии. 1998. Т. 34. № 4. С. 485.
Плетнева Е.В. Особенности моторной асимметрии у крыс с генетической эпилепсией // Журн. высш. нерв. деят. 1999. Т.49. № 3. С. 483.
Плетнева Е.В., Иоффе М.Е. Исследование моторной асимметрии у крыс с аудиогенными судорогами // ХХХ Всерос. cовещ. по пробл. высш. нерв. деят. Т 1. СПб, 2000. С. 371.
Плетнева Е.В., Плескачева М.Г. Исследование моторной асимметрии у крыс линии Крушинского-Молодкиной // ХVIII Всерос. Съезд физиологов, Казань, 2001, 192.
Сташкевич И.С., Воробьева А.Д. Предпочтение конечности при выполнении крысами инструментального навыка: сравнение характера предпочтения на ранних стадиях обучения и в процессе дальнейшей тренировки // Журн. высш. нерв. деят. 1997. Т. 47. № 4. С. 751.
Сташкевич И.С., Куликов М.А. К вопросу о формировании латерализованного двигательного навыка у крыс // Журн. высш. нерв. деят. 2000 . Т. 50. № 3. С. 457.
Сташкевич И.С., Плетнева Е.В., Куликов М.А. Различная устойчивость двигательного предпочтения у крыс к принудительному переобучению // Журн. высш. нерв. деят. 2001. Т. 51. № 6, 694-700.
Bakalkin G.Ya. Neuropeptides induce directional asymmetry in brain and spinal cord: facts and hypotheses // Intern. J. Neurosci. 1989. V. 48. № 1-2. P. 105.
Banerjee P.K., Sneed O.K. Presynaptic Gamma-Hydrobutyric Acid (GHB) and Gamma-Aminobutyric AcidB (GABAB ) receptor mediated release of GABA and glutamate (GLU) in rat ventrobasal nucleus (VB): A possible mechanism for the generation of absence-like seizures induced by GHB // J. Pharmacol. and Exptl. Therap. 1995. V.273. P. 1534.
Barneod P., Van der Loos H. Direction of handedness linked to hereditary asymmetry of a sensory system // Proc. Nat. Acad. Sci. USA. 1993. V. 90. № 8. P. 3246.
Betancur C., Sandi C., Vitiello S. et al. Activity of the hypothalamo-pituitary-adrenal axis in mice selected for left- or right-handedness // Brain Res. 1992. V. 589. P. 302.
Biddle F.G., Coffaro C.M., Zier J.E., Eales B.A. Genetic variation in paw preference (handedness) in the mouse //Genome. 1993. V. 36. № 5. P. 935.
Biddle F.G., Eales B.A. The degree of lateralization of paw usage (handedness) in the mouse is defined by three major phenotypes // Behav Genet. 1996. V. 26. № 4. P. 391.
Biddle F.G., Eales B.A. Mouse genetic model for left-right hand usage: context, direction, norms of reaction, and memory // Genome. 1999. V. 42. № 6. P. 1150.
Bulman-Fleming M.B., Bryden M.P., Rogers T.T. Mouse paw preference: effects of variations in testing protocol // Behav. Brain Res. 1997. V. 86. № 1. P. 79.
Cabib S., D'Amato F.R., Neveu P.J. et al. Paw preference and brain dopamine asymmetries // Neuroscience. 1995.V 64. № 2. P. 427.
Collins R.L. On the inheritance of handedness. I. Laterality in inbred mice // J. Hered. 1968. V. 59. № 1. P. 9.
Collins R.L. On the inheritance of direction and degree of asymmetry. In: Cerebral lateralization in nonhuman species. Ed. Glick S.D. N.Y.: Acad.Press; 1985. P.41.
Collins R.L. When left-handed mice live in right-handed worlds // Science. 1975. V.187. № 4172. P. 181.
Collins R.L. Reimpressed selective breeding for lateralization of handedness in mice // Brain Res. 1991. V. 564. № 2. P. 194.
Denenberg V.H. Hemispheric laterality in animals and the effects of early experience // Behav. Brain Sci. 1981. V. 4. № 1. P. 1.
Denenberg V.H. Evolution proposes and ontogeny disposes // Brain Lang. 2000. V. 73. № 2. P. 274.
Denenberg V.H., Mobraaten L.E., Sherman G.F. et al. Effects of autoimmune uterine/maternal environment on cortical ectopias, behavior and autoimmunity // Brain Res. 1991a.V. 563. № 1. P. 114.
Denenberg V.H., Sherman G.F., Schrott L.M. et al. Spatial learning, discrimination learning, paw preference and neocortical ectopias in two autoimmune strains of mice // Brain Res.1991b. V. 562. № 1. P. 98.
Denenberg V.H., Sherman G.F., Morrison L. et al. Behavior, ectopias and immunity in BD/DB reciprocal crosses // Brain Res. 1992. V. 571. № 2. P. 323.
Depaulis A., Vergnes M., Marescaux C. Endogenous control of epilepsy: the nigral inhibitory system // Progr. Neurobiol. 1994. V. 42. № 1. P. 33.
Deransart C., Marescaux C., Depaulis A. Involvment of nigral glutamatergic inputs in the control of seizures in a genetic model of absence epilepsy in the rat // Neuroscience. 1996.V. 71. № 3. P. 721.
Fride E., Collins R.L., Skolnick P, Arora P.K. Strain-dependent association between immune function and paw preference in mice // Brain Res. 1990. V. 522. № 2. P. 246.
Glick S.D., Jerussi T.P., Zimmerberg B. Behavioral and neuropharmacological correlates of nigrostriatal asymmetry in rats. In: Lateralization in the Nervous System. Eds. Harnad S., Doty R.W., Goldstein L. et al. 1977. N.Y.: Acad.Press. P.213.
Kounin J.S. Laterality in monkeys // J. Genet. Psychol. 1938. V.52. P. 375.
Lehman R.A.W. The handedness in rhesus monkeys // Neuropsychologia. 1970. V.16. № 2. P. 33.
Levitan S., Reggia J.A A computational model of lateralization and asymmetries in cortical maps // Neural Comput. 2000. V. 12. № 9. P. 2037.
Lipp H.P., Collins R.L., Hausheer-Zarmakupi Z. et al. Paw preference and intra-/ infrapyramidal mossy fibers in the hippocampus of the mouse // Behav. Genet. 1996. V. 26. № 4. P. 379.
Martin D., Webster W.G. Paw preference shifts in the rat following forced practice // Physiol. and Behav. 1974. V.13. № 6. P. 745.
McManus I.C. Are paw preference differences in HI and LO mice the result of specific genes or of heterosis and fluctuating asymmetry? // Behav Genet. 1992. V. 22. № 4. P. 435.
Miklyaeva E.I., Ioffe M.E., Kulikov M.A. Innate versus learned factors determining limb preference in the rat // Behav. Brain Res. 1991. V. 46. № 2. P. 103.
Miklyaeva E.I., Bures J. Reversal of handedness in rats is achieved more effectively by training under peripheral than under central blockade of the preferred forepaw // Neurosci. Lett. 1991. V. 125. № 1. P. 89.
Milisen R. The effect of training upon the handedness of the rat in an eating activity // Psychol. Monogr. 1937. V. 49. P. 234.
Neveu P.J., Barneod P., Vitiello S., LeMoal M. Brain modulation of the immune system: association between lymphocyte responsiveness and paw preference in mice // Brain Res. 1988. V. 457. № 2. P. 392.
Neveu P.J., Betancur C., Barneod P. et al. Functional brain asymmetry and murine systemic lupus erythematosis // Brain Res. 1989. V. 498. № 1. P. 159.
Nielsen D.M., Visker K.E., Cunningham M.J. et al. Paw preference, rotation, and dopamine function in Collins HI and LO mouse strains // Physiol. and Behav. 1997.V. 61. № 4. P. 525.
Pediconi M.F., Roccamo de Fernancez A.M., Barrantes F.J. Asymmetric distribution and down-regulation of the muscarinic acetylcholine receptor in rat cerebral cortex // Neurochem. Res. 1993. V. 18. № 5. P. 565.
Peterson G.M Mechanisms of handedness in the rat // Comp. Psychol. Monogr. 1934. V. 9. P. 1.
Peterson G.M. Transfers in handedness in the rat from forced practice // J. Compar. and Physiol. Psychol. 1951. V. 44. № 2. P. 184.
Signore P., Chaoui M., Nosten-Bertrand M. et al. Handedness in mice: comparison across eleven inbred strains // Behav Genet. 1991. V. 21. № 4. P. 421.
Schmidt S.L., Manhaes A.C., de Moraes V.Z. The effects of total and partial callosal agenesis on the development of paw preference performance in the BALB/cCF mouse // Brain Res. 1991.V. 545. № 1-2. P. 123.
Steriade M., Deschenes M. The thalamus as a neuronal oscillator // Brain Res. 1984. V. 320. № 1.P. 1.
Tan U., Kutlu N. The relationships between paw preference and the right- and left-brain weights in male and female adult cats: ipsilateral and contralateral motor control with regard to asymmetric postural and manipulative actions // Int. J. Neurosci. 1993. V. 69. № 1-4. P. 21.
Tsai L.S, Maurer S. «Right-handedness» in white rats // Science. 1930. V. 72. P. 436.
Tsutsui J., Terra V.C., Oliviera J.A.C., Garcia-Cairasco N. Neurioethological evaluation of audiogenic seizures and audiogenic-like seizures by microinjection of bicuculline into the inferior colliculus. I. Effects of midcollicularknife cuts // Behav. Brain Res. 1992. V. 52. № 1. P. 7.
Warren J.M. Handedness and laterality in humans and other animals // Physiol. Psychol. 1980. V. 8 . P. 351.
Waters N.S., Badura L.L., Denenberg V.H. Differential prolactin responsiveness to stress in left- and right-pawed mice // Brain Res. 1996. V. 724. № 1. P. 112.
Waters N.S., Denenberg V.H. Analysis of two measures of paw preference in a large population of inbred mice // Behav. Brain Res., 1994. V. 63. № 2. P. 195.
Wentworth K.L. Some factors determining handedness in the white rat // Genet. Psychol. Monogr. 1942. V. 26. P. 55.
[1] Нередко используется термин «латерализация полушарий», который, как правило, включает все перечисленные виды асимметрии. В настоящей статье речь идет о «моторном предпочтении одной конечности», что является лишь одним из аспектов общей проблемы латерализации.
[2] Как правило, объектом таких исследований являются крысы или мыши (реже кошки), причем первые обычно используются в чисто поведенческих исследованиях, а вторые – в работах по генетике поведения. Данные, полученные на животных одного вида грызунов, часто эктраполируются на другой, хотя, конечно, не стоит забывать о межвидовых различиях. Особую группу составляют работы на обезьянах, целью которых в значительной степени является выяснение филогенетических основ предпочтения конечности у человека.