ЦИТОАРХИТЕКТОНИЧЕСКАЯ АСИММЕТРИЯ КОРКОВЫХ ПОЛЕЙ И ХВОСТАТОГО ЯДРА МОЗГА ЧЕЛОВЕКА
И.Н. Боголепова, Л.И. Малофеева, Н.С. Оржеховская, Т.В. Белогрудь
Функциональная неравнозначность полушарий мозга известна со времен Брока [25].
Согласно современной концепции о структурно-функциональной асимметрии мозга, сложившейся в результате многочисленных клинических и экспериментальных исследований, существуют значительные различия функций левого и правого полушарий мозга человека, их функциональная неидентичность в процессе формирования второй сигнальной системы и в обеспечении речи [13, 14, 15, 30].
Левое полушарие связано преимущественно с реализацией специфически человеческих речевых функций, а также с аналитической абстрактной деятельностью на речевой и знаковой основе. Оно участвует в прогнозировании сложных моторных и психических актов. Правое полушарие является в основном ведущим в восприятии пространства, времени и имеет особое значение в организации эмоциональной жизни [16, 23].
Выявлены различия нервных и психических расстройств при поражении правого и левого полушария [9, 1].
Для понимания механизмов функциональной асимметрии мозга, его сложных психических, речевых и двигательных функций большое значение имеет исследование особенностей структурной организации коры и подкорковых образований мозга человека в левом и правом полушарии.
В литературе имеются лишь единичные работы, посвященные данному вопросу [10, 11, 12, 17, 20], выполненные традиционными архитектоническими методами и касающиеся в основном особенностей строения борозд, извилин и отдельных корковых полей.
Целью настоящей работы было выявление в левом и правом полушарии мозга человека особенностей строения корковых полей, ответственных за обеспечение сложных психических, речевых и двигательных функций, а также структурной организации подкорковых образований.
Были изучены речедвигательные поля (44 и 45); корковые поля двигательного анализатора (4 и 6); поле 43, играющее непосредственную роль в звукопроизнесении; поле 8, принимающее участие в осуществлении пространственной ориентации и регуляции произвольных двигательных актов; поля 47 и 46, чисто лобные формации, принимающие участие в регуляции программирования и осуществления сложных интеллектуальных процессов; поля 39 и 40 нижнетеменной области, связанные с процессами чтения, письма, рисования, формирования целенаправленных действий; хвостатое ядро, участвующее в управлении двигательными механизмами и в организации сложных форм поведения.
Исследование проводили на 10 полушариях мозга взрослых людей в возрасте от 30 до 60 лет. Изучали серии фронтальных парафиновых срезов толщиной 20 мкм, окрашенных крезилом фиолетовым. Архитектонические структуры исследовали на качественном и количественном уровне. С помощью телевизионно-оптических устройств “TAS” и “Видеоплан,” сопряженных с микроскопом, проводили измерение суммарной объемной фракции нейронов и глии и площади профильных полей пирамидных нейронов в слоях III и V корковых полей, а также в подкорковых структурах. Измерения в левом и правом полушариях мозга проводили на вершине извилины в идентичных по топографии местах. В каждом полушарии измерялось 100 нейронов, исследовалось 50 полей зрения. Составлены цитоархитектонические карты корковых полей в правом и левом полушарии. Распределение нейронов на классы проводилось в соответствии с классификацией, разработанной Малофеевой Л.И и Оржеховской Н.С. Выделено 5 классов нейронов: сверхмалые (до 70,0 мкм2), малые (70,1-170,0 мкм2), средние (170,1-290,0 мкм2), крупные (290,1-500,0 мкм2), сверхкрупные (свыше 500,1 мкм2). Коэффициент асимметрии средней величины профильного поля нейронов (Мпп) в левом и правом полушариях мозга определялся по формуле: [Mпп(лев)–Мпп(прав)]/0.5[Мпп(лев)+Мпп(прав)]*100%. Была проведена статистическая обработка результатов: достоверные различия считались значимыми при Р³0,05.
Сведения о подобном изучении структурной организации корковых и подкорковых образований мозга человека в левом и правом полушарии с применением количественного анализа и электронно-вычислительной техники отсутствуют.
В результате исследований речедвигательных полей выявлено четкое различие правого и левого полушария мозга человека по максимальным, средним и минимальным величинам площади профильных полей пирамидных нейронов в слоях III и V этих структур. В слое III поля 44 во всех наблюдениях в левом полушарии величина площади профильных полей пирамидных нейронов, равная 287±15, 285±13, 269±19 и 306±13 мкм2, превышала таковую в правом полушарии – 231±13, 207±10, 159±11 и 217±11 мкм2. Эта же закономерность была установлена и при изучении слоя V полей 44 и 45. Так, в слое V поля 44 в левых полушариях величины площади профильных полей пирамидных нейронов, равные 259±10, 265±11, 263±13 и 238±8 мкм2, превышали таковые в правых полушариях – 206±10, 207±9, 221±10 и 206±7 мкм2.
В левом полушарии в поле 44 максимальная величина площади профильных полей пирамидных нейронов в слое III была равна в среднем 525 мкм2, а в правом – 399 мкм2. Та же закономерность была установлена при изучении поля 45. Максимальная величина площади профильных полей пирамидных нейронов в слое III в левом полушарии равнялась в среднем 411 мкм2, в правом – 316 мкм2.
Анализ полей 4 и 6 показал аналогичные структурные особенности их строения в левом и правом полушарии мозга человека. Так, в слое III поля 6 в левых полушариях величина площади профильных полей пирамидных нейронов, равная 302±20, 274±19, 264±18 и 269±14 мкм2, превышала таковую в правых полушариях – 216± 13, 204± 11, 248± 14 и 212± 13 мкм2. В слое V поля 6 величина площади профильных полей пирамидных нейронов в левых полушариях также была больше, чем в правых полушариях.
Эта же закономерность была прослежена при сопоставлении площади профильных полей пирамидных нейронов в слое III и V поля 4 в левом и правом полушарии мозга человека, причем наибольшие различия были выявлены при исследовании клеток Беца.
Таким образом, была выявлена левополушарная доминантность речедвигательных и двигательных корковых полей мозга человека, что коррелирует с левополушарной латерализацией речедвигательных и двигательных функций человека [2, 3, 4, 5, 6].
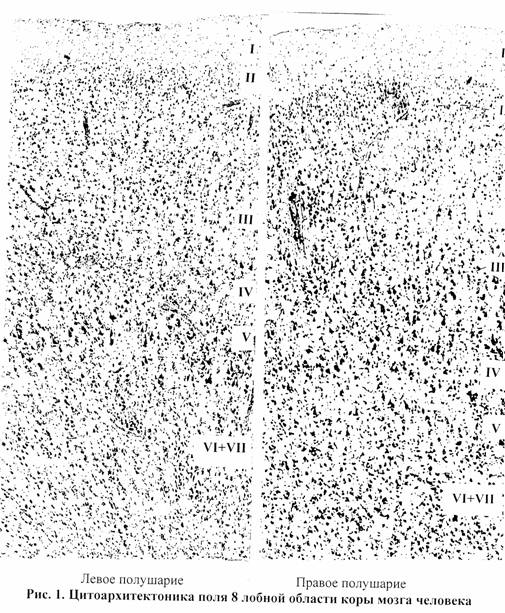
При изучении полей 43, 8, 46, 47 обнаружились определенные различия не только в картине расположения полей по поверхности полушарий, но и в структурной организации указанных образований. Так, например, при сравнении цитоархитектоники поля 8 в правом и левом полушариях обнаруживается, что его фактура слева – четче и определеннее: стратификация коры яснее; радиальная исчерченность, особенно в нижнем этаже коры – выраженнее, границы гранулярных слоев III и V – резче; напротив, справа – некоторая размытость структуры (рис.1).
Вместе с тем, количественный анализ некоторых параметров не выявил устойчивой левой или правополушарной доминантности. Например, достоверность величин поверхности названных выше лобных полей в одних случаях преобладала в левых полушариях,в других – в правых, в третьих – отсутствовала (рис. 2).
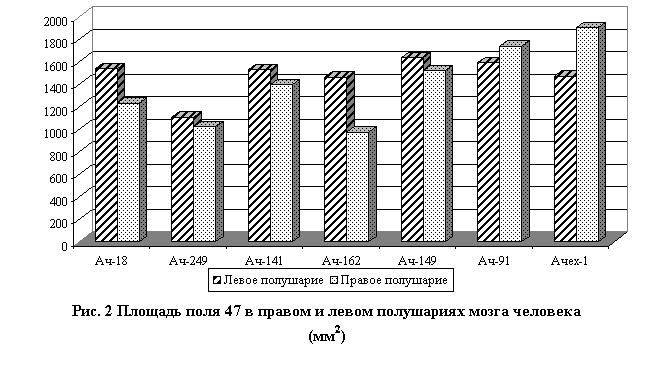
Измерение ширины поперечника коры в этих же лобных структурах также не обнаружило постоянной доминантности какого-либо полушария, так как превалирование данного параметра отмечалось то справа, то слева. Аналогичная картина наблюдалась при изучении отдельных слоев в полях 8, 43, 46, 47.
При исследовании указанных выше полей проявились определенные полушарные различия в размерах пирамидных нейронов и их плотности. Однако, как показала статистическая обработка, если эти различия отчетливо проступали в поле 47 (слой III), то в поле 46 (слой III) они весьма невелики. Так, величина профильного поля 46 (слой III) лишь в незначительном числе случаев оказывалась больше справа - 176±12 мкм2(пр) и 122±8 мкм2(лев), а в поле 47 (слой III) в большинстве случаев - слева.
Также не выявлено достоверных различий между полушариями по величине профильного поля нейронов в слое V полей 46 и 47. В поле 46 лишь в одном случае отмечено достоверное увеличение размеров нейронов в правом полушарии 183±12 (пр) и 124±7(лев). В поле 47 в одном случае наблюдалось абсолютное равенство - 137±8 (пр) и 137±10 (лев).
В полях 8 и 43 в двух наблюдениях из четырех обнаружено преобладание левого полушария, а в двух случаях – правого: так, в слое III поля 8 величина профильного поля пирамидных нейронов равнялась 171±8,8(лев) и 150±10,3(пр); 190±11,7(лев) и 178±11,5(пр) и ,соответственно, - 118±7(пр) и 136±7(лев); 132±6,3(пр) и 167±5(лев) мкм2. Детальное изучение объемной фракции нейронов и глии в слое III и V корковых полей 8, 46, 43, 47 показало значительные колебания этого показателя в левых и правых полушариях и отсутствие его постоянного преобладания в одном из полушарий.
Анализ плотности нейронов в отдельных корковых слоях вышеназванных лобных полей также не выявил повторяющегося превалирования этого показателя в одних и тех же полушариях.
Таким образом, по некоторым цитоархитектоническим параметрам в структурной организации лобных полей 8, 43, 46, 47 отсутствует устойчивое доминирование одного полушария над другим, что подтверждается данными литературы.
Исследование нижнетеменной области коры мозга человека показало, что как в поле 39, так и в поле 40 в слое III3 в изученных случаях ни левое , ни правое полушария достоверно не выявляют устойчивой доминантности по среднему значению профильного поля пирамидных нейронов. Например, в поле 40 в слое III3 среднее значение профильного поля пирамидных нейронов было равно в одном случае – 176,4±4,1 мкм2 слева и 188,6±8,7 мкм2 справа, в другом -–соответственно 159±6,7 и 186,9±11,7 мкм2, в третьем – 216,5±12,3 и 199±8,6 мкм2. В поле 39 в слое III3 в двух случаях из десяти размер нейронов был достоверно больше в левом полушарии, в двух – в правом, в шести случаях достоверно не отличался в обоих полушариях. Так, например, среднее значение площади профильного поля нейронов в слое III3 в мозге Ar-18 в левом полушарии было равно 204,3±5,9 мкм2, в правом – 168,7±5,9 мкм2; в мозге Ан-3 – 214,0±10,3 в левом полушарии и 260,5±17 в правом; а в мозге А-247 – 210,4±10,3 мкм2 и 215,2±6,0 мкм2 соответственно.
Такая же закономерность была выявлена при изучении пирамидных нейронов слоя V. В обоих полях нижнетеменной области они достоверно не отличались по среднему значению профильного поля пирамидных нейронов в левом и правом полушарии в большинстве изученных экземплярах мозга. В одном случае наблюдалось достоверное увеличение среднего значения площади профильного поля нейронов слоя V в правом полушарии (166,5± 8,8мкм2 и 192,3±11,9 мкм2) и в двух случаях в левом полушарии: Ar-40 –218,3± 9,3 (лев) и 163,4±9,6 мкм2 (пр); А-91 – 204,7±8,6 (лев) и 142,4±7,3 мкм2 (пр).
Таким образом, изучение цитоархитектоники полей 39 и 40 нижнетеменной области коры мозга человека в большинстве случаев не выявило преобладания левого или правого полушария мозга по величине площади профильного поля нейронов слоев III и V.
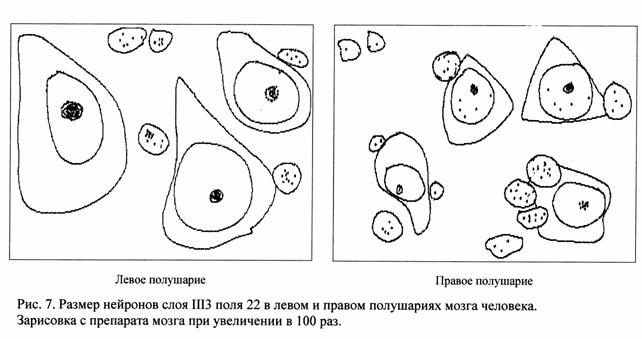
Цитоархитектоническое изучение полей 41 и 22 височной области мозга человека показали особенности топографии и нейронного состава этих полей в левом и правом полушарии. В левом полушарии в поле 22 радиарная исчерченность более тонкая, чем в правом полушарии и лучше выражена в большинстве изученных случаев. Слои коры достаточно четко отграничиваются друг от друга, как в правом, так и в левом полушарии мозга. Однако, имеется некоторая тенденция к более выраженным границам между слоями в левом полушарии по сравнению с правым, указывая тем самым на лучшую горизонтальную исчерченность коры поля 22 в левом полушарии. Ширина слоев III3, V больше в левом полушарии, чем в правом. Слой III3 поля 22 имеет более упорядоченную структуру, по сравнению с правым полушарием. Пирамидные нейроны этого слоя располагаются колонками, что указывает на более высокую степень вертикальной организации поля 22 в левом полушарии, чем в правом (рис.3).

Количественное изучение площади профильного поля нейронов и процентного соотношения отдельных классов нейронов в полях 41 и 22 височной области мозга человека в левом и правом полушарии выявили определенные интересные закономерности. В поле 41 в слое III3 площадь профильного поля нейронов в большинстве случаев оказалась ниже в левом полушарии, чем в правом (рис. 4 и 5).
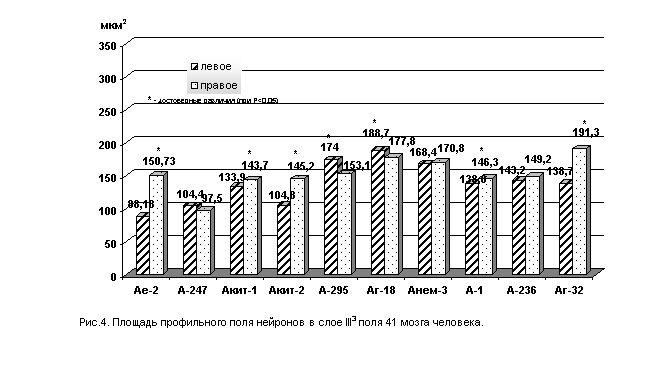
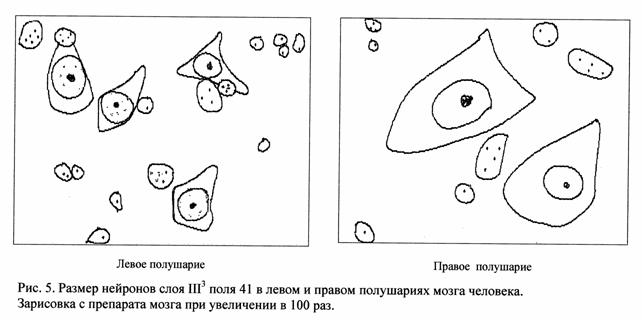
Однако, разница между средним значением площади нервных клеток в обоих полушариях невелика. Так, например, коэффициент асимметрии профильного поля нейронов в слое III3 поля 41 равен: в мозге А-236 - 4,1%, в мозге А-247– 3,4%.
Иная картина наблюдается в поле 22. В слое III3 поля 22 во всех изученных случаях площадь профильного поля нейронов больше в левом полушарии по сравнению с правым (рис. 6 и 7). Коэффициент асимметрии по данному показателю оказался выше, чем в поле 41. Так, коэффициент асимметрии профильного поля нейронов в слое III3 поля 22 равен: в мозге А-236 - 44,1%, в мозге А-247 – 6,6%.
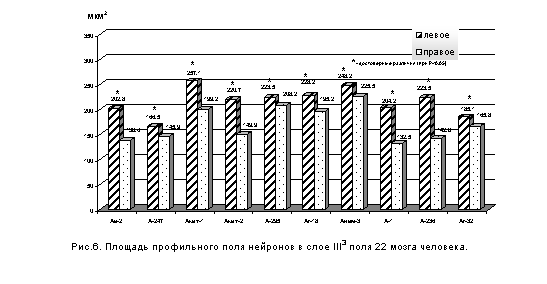
При сравнении средних величин профильного поля нейронов слоя III3 в полях 41 и 22 было выявлено, что в левом полушарии размер нейронов поля 22 превышает аналогичный показатель для поля 41 во всех изученных мозгах. В правом полушарии площадь профильного поля нейронов остается выше в поле 22 лишь в половине случаев.
Анализ классового состава нейронов по величине профильного поля нейронов в слое III3 выявил отличия между полем 41 и 22, а также между левым и правым полушариями. В поле 41 в большинстве изученных случаев процент нейронов малого размера оказался выше в левом полушарии, чем в правом, и составил до 82%, тогда как в правом полушарии этот показатель равен 64-75%. В правом полушарии в поле 41 увеличивается процент нейронов среднего размера (до 29%) и крупного размера (до 7%). В поле 22 наблюдается противоположная картина. Процент нейронов среднего и крупного размера больше в левом полушарии, чем в правом (рис. 8).

Среднее значение площади профильного поля нейронов в этом поле соответственно выше в левом полушарии по сравнению с правым.
Результаты, полученные при исследовании площади профильного поля нейронов в полях 41 и 22 в левом и правом полушарии мозга человека позволяют сделать вывод о том, что разные по функциональному значению корковые слуховые формации имеют разный цитоархитектонический рисунок, характеризующийся в частности, разной величиной площади профильного поля нейронов.
В литературе высказывается положение о том, что специфические лингвистические функции связаны с ассоциативными полями, расположенными в левом полушарии [24]. Известно, что в левом полушарии более эффективно осуществляется обработка вербальных стимулов речевого происхождения, и интенсивнее происходит обработка сложных грамматических и фонематических конструкций [7, 8, 18]. Действительно, поле 22, являясь проекционно-ассоциативным, в левом полушарии содержит более крупные нейроны, чем в правом. Левополушарную асимметрию площади профильного поля нейронов поля 22 также можно объяснить тем, что поле 22 (зона Вернике) имеет тесные связи с речедвигательной зоной коры (с полем 45) мозга, функциональная активность которой, как известно, более сильно выражена в левом, доминантном по речи, полушарии. Кроме того, показана определенная связь между ведущей рукой и латерализацией слуховой функции у человека. Так, функция правого уха более латерализована у правшей, характеризующихся выраженной левополушарной асимметрией речевоспринимающей функции мозга [26, 27, 28].
Поле 41 является проекционным и содержит нейроны малого размера как в левом, так и в правом полушарии мозга. Оно относится к первичной слуховой коре и представляет собой корковый конец слухового анализатора, куда поступает звуковая информация и где происходит ее первичная кодировка. При несильно выраженной асимметрии величины профильного поля нейронов поля 41, можно лишь говорить о тенденции доминирования правого полушария по данному признаку, поскольку величина среднего значения площади профильного поля нейронов в правом полушарии была несколько выше, чем в левом. Данное наблюдение, по-видимому, связано с тем, что в процессе первичной кодировки звуковых стимулов правое полушарие, в большей степени, чем левое, играет роль своеобразного «фильтра», выделяя «сигнал» из «шума» [8].
По сравнению с полем 41, в поле 22 нейроны имеют более крупные размеры, что отражает большую функциональную специфичность поля 22 как ассоциативного звена в сложном процессе восприятия и обработки звуковых стимулов различной природы [18]. Более выраженная асимметрия величины площади профильного поля нейронов в поле 22 по сравнению с полем 41, по-видимому, связана с тем, что поле 22 является филогенетически более новым образованием коры мозга человека и сложность строения и функционирования этой структуры связана, прежде всего, с появлением такого нейролингвистического феномена как речь, характерной только для человека.
Таким образом, результаты настоящего исследования показали значительные отличия между величиной профильного поля нейронов в поле 41 и 22 в левом и правом полушарии мозга человека. Поле 22 характеризуется большей величиной профильного поля нейронов и левополушарной асимметрией по изученному признаку. Поле 41 характеризуется меньшей величиной профильного поля нейронов и тенденцией к правополушарной асимметрии по данному показателю. Степень асимметрии величины площади профильного поля нейронов в большей степени выражена в поле 22, по сравнению с полем 41 во всех изученных случаях.
Сравнительный анализ структурной организации хвостатого ядра справа и слева в каждом случае не выявил преобладания какого-либо полушария по исследуемым параметрам: в ряде наблюдений доминировало левое полушарие, в ряде наблюдений – правое (рис. 9).
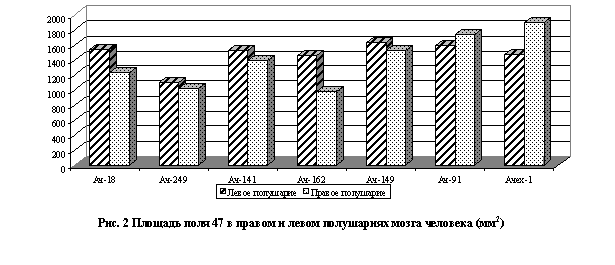
Таким образом, настоящая работа показывает особенности структурной организации ряда корковых и подкорковых образований в левом и правом полушарии мозга человека. Полученные нами данные свидетельствуют о наличии некоторых морфологических признаков их асимметрии. Результаты исследований подтверждают концепцию О.С. Адрианова (1979), считающего, что на уровне коры большого мозга человека «не существует какого-либо универсального показателя или критерия функциональной асимметрии».
Исследование структурной асимметрии мозга человека тесно связано с изучением индивидуальной вариабельности мозга. Психологи, клиницисты, нейроморфологи и физиологи уделяют большое внимание индивидуальной вариабельности и типологическим особенностям строения и функции мозга.
В работах Е.П. Кононовой [12] и других [21, 22, 19, 10] были показаны определенные индивидуальные особенности расположения и количества борозд, топографии и площади поверхности отдельных корковых полей мозга человека. Однако морфологические критерии индивидуальных и типологических особенностей коры и подкорковых образований мозга взрослого человека до настоящего времени окончательно не вскрыты. Для этого необходимы исследования с применением количественного анализа и электронно-вычислительной техники.
В связи с вышеуказанным, следующая задача нашей работы заключалась в исследовании вариабельности мозга человека и в поиске морфологических критериев, характеризующих особенности архитектонических формаций у разных индивидуумов. Предполагалось выяснить, какие цитоархитектонические признаки являются основными для характеристики структурной организации мозга, только ли в коре могут быть выявлены специфические особенности строения мозга у разных индивидуумов и как уровень структурной асимметрии корковых формаций мозга человека коррелирует с индивидуальной вариабельностью.
В результате проведенного исследования были получены факты, свидетельствующие о наличии некоторых морфологических признаков индивидуальной вариабельности мозга человека. Так, при сравнении индивидуальной вариабельности строения поля 8 в левых полушариях были установлены следующие различия в величинах суммарной объемной фракции нейронов и глии. В большинстве изученных случаях суммарные объемные фракции нейронов и глии достоверно отличались друг от друга (0,122±0,006; 0,083±0,03; 0,061±0,004). В правых полушариях эта разница в поле 8 выражена отчетливее. Все правые полушария резко отличаются друг от друга (0,135±0,005; 0,115±0,005;0,076±0,004; 0,141±0,004). При исследовании суммарной объемной фракции нейронов и глии в поле 43 были выявлены достоверные различия также в большинстве случаев в правых полушариях (0,106±0,005; 0,083±0,005; 0,117±0,005).
Такая же закономерность была обнаружена и в поле 44, где как в левых, так и в правых полушариях мозга у разных индивидуумов имелись достоверные различия суммарной объемной фракции нейронов и глии (слева – 0,16±0,01; 0,13±0,01; 0,14±0,01 и справа – 0,15±0,01; 0,12±0,01; 0,10±0,01; 0,18±0,01).
Исследование площади профильных полей пирамидных нейронов слоев III и V выявило определенные различия и этих показателей в корковых полях у разных индивидуумов. Так, в поле 8 в левом полушарии средняя величина профильного поля пирамидных нейронов слоя III колебалась в пределах от 136±6 мкм2 до 190±11 мкм2. В правых полушариях различия величины профильного поля нейронов были меньше, чем в левых полушариях.
В поле 39 среднее значение профильного поля пирамидных нейронов слоя III в левых полушариях варьировало незначительно от 204 мкм2 до 238,1 мкм2. Достоверных различий по данному признаку в большинстве исследованных случаев между левыми полушариями не наблюдалось. В правых полушариях отмечалась большая вариабельность среднего значения профильного поля нейронов: от 168 мкм2 до 269,2 мкм2. В слое V этого поля в левых полушариях профильные поля нейронов варьировали от 168,5 мкм2 до 208,3 мкм2 , а в правых – от 178,9 мкм2 до 215,1 мкм2. Как в левых, так и в правых полушариях в большинстве изученных случаев в слое V поля 39 отличия были недостоверными.
В поле 40 в слое III в левых полушариях площадь профильного поля нейронов варьировала от 159,2 мкм2 до 244,3 мкм2. В правых полушариях она колебалась в меньших пределах - от 164,8 мкм2 до 199,9 мкм2. Такая же закономерность выявлена в слое V этого поля. В левых полушариях площадь профильного поля нейронов варьировала от 159,9 мкм2 до 240,7 мкм2 , а в правых – от 162,8 мкм2 до 213,2 мкм2.
Изучение средних величин площади профильного поля пирамидных нейронов слоев III и V поля 44 мозга у разных индивидуумов также обнаружило определенные колебания этих показателей. Так, в левых полушариях средняя величина площади профильного поля в слое III колебалась от 269±19 мкм2 до 306±13 мкм2, а в правых – от 159±11 мкм2 до 231±13 мкм2.
Итак, в результате проведенных измерений установлено, что корковые поля мозга человека отличаются у разных индивидуумов по среднему значению площади профильного поля пирамидных нейронов в слоях III и V . Однако эта особенность в различных корковых полях проявляется по-разному. В большей степени она выражена в лобных полях и в меньшей- в полях нижнетеменной области.
Индивидуальная вариабельность строения изученных архитектонических формаций выявилась также и в распределении нервных клеток слоев III и V по их размерам. Исследование площади профильного поля корковых полей позволило разделить клетки по их размерам на пять классов: 1-й класс- сверхмалые нейроны (до 45 мкм2), 2-й класс- малые (от 46 до 90 мкм2), 3-й класс – средние (от 91 до 270 мкм2), 4-й класс – крупные (от 271 до 500 мкм2), 5-й класс – сверхкрупные (свыше 500 мкм2). Для корковых образований характерно преобладание нейронов 2-го и 3-го классов, которые по степени вариабельности значительно уступали классу сверхмалых нейронов. Так, например, в левых полушариях поля 8 количество клеток 1-го класса колебалось в пределах 4-12%, 2-го класса – в пределах 16-36%, а 3-го класса – 52-80%.
Сопоставление гистограмм профильного поля нейронов в слоях III и V полей 39 и 40 в левых и правых полушариях показало, что в большинстве случаев, даже там, где не наблюдалось отличий по среднему значению площади профильного поля, отмечались индивидуальные различия в процентном составе нейронов по классам от мозга к мозгу, а также в левом и правом полушариях одного мозга. Наибольшие различия наблюдались в слое III поля 40 в левых полушариях по классам средних и крупных нейронов.
Изучение хвостатого ядра также обнаружило морфологические критерии, указывающие на особенности строения этого образования у разных индивидуумов. В хвостатом ядре в правых полушариях по сравнению с левыми вариации проявлялись резче: в трех случаях из четырех достоверно различались между собой по площади профильного поля (56±2 мкм2; 74±4 мкм2; 57±3 мкм2; 105±5 мкм2) и по суммарной объемной фракции нейронов и глии (0,059±0,003; 0,089±0,003; 0,037±0,002; 0,069±0,003). Наибольший процент от общего количества нейронов составляли клетки 1-го и 2-го классов, а наименьшей вариабельностью отличались клетки 3-го класса. Сходная картина была выявлена при изучении плотности нейронов этого образования.
Таким образом, проведенное исследование выявило, что площадь профильного поля нейронов, суммарная фракция нейронов и глии и распределение нейронов по классам могут служить морфологическими критериями, свидетельствующими об индивидуальных особенностях структурной организации корковых и подкорковых образований мозга человека. Диапазон колебаний указанных показателей различен в отдельных корковых полях. Так, процентное содержание пирамидных нейронов средней величины в поле 43 колебалось в пределах от 32% до 64% у разных индивидуумов, а в поле 8 – от 52% до 80%. Эти данные свидетельствуют о возможных функциональных особенностях корковых полей у этих лиц.
При сопоставлении структурной организации архитектонических формаций на корковом и подкорковом уровнях обнаружена различная картина нейронного состава. Так, для полей 43 и 8 характерно преимущественно преобладание малых и средних пирамидных нейронов, для хвостатого ядра- средних и крупных. Это говорит о том, что по-видимому, морфологические критерии индивидуальной вариабельности у корковых и подкорковых структур могут принципиально отличаться.
Для характеристики морфологических критериев важно, что степень выраженности индивидуальной вариабельности мозга человека по-разному проявляется не только в различных корковых полях и подкорковых структурах, но в разных полушариях. Можно предположить, что различный уровень структурной асимметрии цитоархитектонических формаций мозга человека в определенной мере определяет его индивидуальную вариабельность и индивидуальные когнитивные способности.
1. Белый Б.И. Психопатология очагового поражения лобных долей в свете функциональной асимметрии мозга. В кн.: Функциональная асимметрия и адаптация человека. М., 1976, с.205-207.
2. Bogolepova I.N., Uylings H., Malofeeva L.I., Amunts K., Zilles K. Broca’s language area from a neuroanatomical and developmental perspective. In book “The Neurocognition of Language,” 1999, Oxford, Ch. 10, pp. 320–336.
3. Боголепова И.Н., Малофеева Л.И., Улингс Х.Б.М. Структурная асимметрия речедвигательных полей 44 и 45 коры мозга человека в постнатальном онтогенезе. Ж. «Бюллетень экспериментальной биологии и медицины», 1999, №10, с. 471-476.
4. Боголепова И.Н., Малофеева П.И., Белогрудь Т.В., Ванина А.Н. Глио-нейрональные соотношения в речедвигательных полях 44 и 45 в левом и правом полушариях мозга человека в раннем постнатальном онтогенезе. Ж. “Российские морфологические ведомости”, 1999, .№3-4, с. 187–191.
5. Боголепова И.Н., Малофеева П.И., Белогрудь Т.В., Улингс Х. Структурная асимметрия речедвигательных полей 44 и 45 коры мозга человека в постнатальном онтогенезе. Ж. «Бюллетень экспериментальной биологии и медицины», 1999, т.128, №10, с.471–475.
6. Боголепова И.Н. Предпосылки структурной асимметрии речедвигательной зоны Брока мозга человека. Ж. “Вестник Российской Академии медицинских наук”, 2001, №4, с. 61–64.
7. Вартанян ИА. Звук-слух-речь. Ленинград: Наука, 1981.
8. Вартанян И.А. Шмигидина Г.Н. Слуховая кора. В кн. Слуховая система: 1990, с.259.
9. Доброхотова Т.А., Брагина Н.Н. Функциональная асимметрия человека. М.: Медицина, 1981, 288 с.
10. Зворыкин В.П. Нейроморфологическое свидетельство индивидуальных различий в зрении человека. Ж. «Архив анат., гистол. и эмбриол.», 1981, т.81, в.10, с.21-24.
11. Кононова Е.П. Вариабельность строения коры большого мозга. Нижняя фронтальная извилина взрослого человека. Труды ин-та мозга, 1935, в.1, с.49-115.
12. Кононова Е.П. Вариабельность строения коры большого мозга. Лобная область взрослого человека. Труды ин-та мозга, 1938, в. 3-4, с.213-274.
13. Лурия А.Р. Мозг человека и психические процессы, М., 1970, 419 с.
14. Лурия А.Р. Основы нейропсихологии. М. Изд-во МГУ, 1973, 388 с.
15. Милнер П. Физиологическая психология. М., 1973.
16. Мосидзе В.М., Акбардия К.К. Функциональная асимметрия и асимметрия полушарий мозга. Тбилиси, 1973, 84 с.
17. Преображенская Н.Ч. Постнатальное развитие затылочноцй области мозга человека. Труды ин-та мозга, 1948, т.6, с.44-58.
18. Радионова ЕА. Анализ звуковых сигналов в слуховой системе. Ленинград: Издательство "Наука", 1987.
19. Саркисов С.А. Индивидуальная вариабельность структурных особенностей коры мозга человека. В кн.: Структурные основы деятельности мозга. М. «Медицина», 1980, с.27-34.
20. Соколовская Н.Ю. О методе оценки степени вертикальной организации структур коры большого мозга. Ж. «Архив анат. гистол. и эмбриол.», 1976, т.LXX, в. 6, с. 105-109.
21. Станкевич И.А., Шевченко Ю.Г. Вариабельность строения коры большого мозга. Нижнепариетальная область у взрослого человека. Труды ин-та мозга, М., 1935, т.1, в.1, с.119-172.
22. Филимонов И.Н. Вариабельность строения коры большого мозга человека. В кн.: 2-я сессия нейрохирургического совета. М.-Л., 1938, с.111-117.
23. Шмидт Е.В., Бабенкова С.В. Функциональная асимметрия головного мозга человека. Ж. «Невропатол. и психиатр.». 1975, N12, с.1767-1775.
24. Binder JR, Frost JA, Hammeke TA, Rao SM, Cox RW. Function of the left planum temporale in auditory and linguistic processing. Brain, 1996, 119 (Pt 4): 1239-47.
25. Broca P. Sur la facilite du language articule. Bull. Soc. Antropol. (Paris), 1865,v.6, p.377-393.
26. Khalfa S, Veuillet E, Collet L. Influence of handedness on peripheral auditory asymmetry. Eur J Neurosci, 1998, 10 (8), р.2731-2737.
27. Kimura D. Cerebral dominance and the perception of verbal stimuli. Canad. J. Psychol., 1961, 15 (№1), р. 166-171,.
28. Koff E, Naeser MA, Pieniadz JM, Foundas AL, Levine HL. Computed tomographic scan hemispheric asymmetries in right- and left-handed male and female subjects. Arch Neurol., 1986, 43 (5), р.487-491.
29. McFadden D. A speculation about the parallel ear asymmetries and sex differences in hearing sensitivity and otoacoustic emissions. Hear Res, 1993,68 (2), р. 143-151.
30. Sperry R.W. Cerebral organization and behavior. Science, 1961, v.133, p.13-22.