О СТРУКТУРНОЙ ОБУСЛОВЛЕННОСТИ ФУНКЦИОНАЛЬНОЙ МЕЖПОЛУШАРНОЙ АСИММЕТРИИ МОЗГА
С.В. Алексеенко
Институт физиологии им. И. П. Павлова РАН
Многочисленные экспериментальные и клинические данные (Л.И. Леушина с соавт., 1982; Я.А. Меерсон, 1989; А.А. Невская, Л.И. Леушина, 1990; S. Christman, 1989) свидетельствуют о различиях в способах переработки информации в левом и правом полушариях головного мозга. Естественно было бы предположить, что эти различия отражены и в структурной организации полушарий. Однако аксиоматическое утверждение о взаимообусловленности структуры и функции в применении к дихотомии мозга к настоящему времени не имеет достаточного экспериментального подтверждения. Это объясняется, с одной стороны, сложностью процессов восприятия, памяти и распознавания, основные этапы которых еще до конца не выяснены и не локализованы, с другой стороны, несовершенством имеющихся к настоящему времени методов исследования. Целью данной работы является не столько описание имеющихся немногочисленных данных о структурной асимметрии мозга, сколько привлечение внимания к изучению этой проблемы.
У человека структурные различия полушарий головного мозга к настоящему времени выявлены только в корковых полях высших уровней, связанных с речевой деятельностью и пространственной ориентацией. В этих областях наблюдается неодинаковое строение борозд и извилин, отличается форма и площадь корковых полей 22, 39, 44, 45, 47, плотность и размер нейронов отдельных цитоархитектонических слоев (Е.П. Кононова, 1935; О.С. Адрианов, 1999; N. Geschwind, W. Levitsky, 1968; A.M. Galaburda et al., 1978; И.Н. Боголепова, 1981, 2001). Эти различия полей по базовым морфологическим показателям могут служить доказательством особой биологической значимости межполушарной асимметрии на этом корковом уровне.
У животных асимметричное функционирование полушарий головного мозга в течение многих лет подвергалось сомнению. Считалось, что функциональная межполушарная асимметрия (ФМА) является “прежде всего достоянием эволюции гоминид, особенно человека, и возникла в результате становления социального характера его деятельности и поведения” (О.С. Адрианов, 1999). Однако в экспериментальных исследованиях были получены доказательства существования у животных латеральной специализации больших полушарий головного мозга (В.Л. Бианки, 1985, 1989; Н.В. Праздникова с соавт. 1997, 1999). Причем, ФМА у животных была выявлена не только в ассоциативных, но и в проекционных полях коры. Показано, что преобладание одного из полушарий различается в полях разных сенсорных модальностей, степень выраженности асимметрии меняется при разных условиях стимуляции, зависит от типа решаемой животным задачи (В.Л. Бианки, 1985). Это свидетельствует о том, что у животных преимущество левого полушария в процессе реализации одних элементарных функций может сочетаться с преимуществом правого полушария в выполнении других. Однако, на уровне реакций отдельных клеток межполушарные различия не очевидны (D.H. Hubel, T.N. Wiesel, 1962; G.A. Orban, 1984). Из многочисленных нейрофизиологических исследований активности клеток разных зрительных полей коры кошки только в поле 21а выявлены различия в размере рецептивных полей и степени выраженности некоторых функциональных свойств нейронов разных полушарий (И.М. Габибов, В.Д. Глезер, 1985; А.В. Болотов, 2001).
К настоящему времени имеются сведения о структурных различиях полушарий мозга у разных животных: асимметричном расположении борозд в височной области у кошек и обезьян, превосходстве по размеру и весу правого полушария у мышей, крыс, кроликов и кошек, большей толщине коры правого полушария у крыс (В.Л. Бианки, 1985, 1989) , неодинаковом количестве нейронов в первичных корковых полях у крыс (G.D. Rosen, 1996). Очевидно, что этих данных недостаточно для утверждения, что в основе функциональной латерализации полушарий лежит структурная асимметрия. При этом отметим неправомерность прямых сравнений структурных и функциональных особенностей отделов мозга у животных разного уровня организации, даже если есть показания о гомологическом соответствии этих отделов. В качестве примеров можно рассмотреть формирование избирательных свойств зрительных нейронов. Известно (А.Я. Супин, 1981), что в ретино-геникуло-корковом пути нейроны, избирательные к направлению движения, представлены у кролика и суслика в сетчатке, а у кошки и обезьяны такие клетки обнаружены только в коре. Неспециализированные концентрически организованные рецептивные поля у кошки встречаются только в подкорковых структурах, а у обезьяны, кроме того, и в первичном корковом поле V1.
Известно, что обнаружение, различение и опознавание сигналов из внешней среды и управление поведением организма – многоэтапный процесс. Для его реализации в головном мозге имеется много структурных уровней. До настоящего времени еще неясно насколько перекрываются нейроанатомические субстраты этих процессов. Только в корковом отделе головного мозга выделяют 10-14 иерархических уровней полей, связанных с восприятием и обработкой зрительной информации (около 32 полей у обезьяны (D.C. Van Essen et al., 1992) и около 22 полей у кошки (J.W. Scannel et al., 1995). Для выявления этой иерархии были использованы морфологические критерии - определенная послойная локализация тел инициальных клеток и их аксонных окончаний в связях между полями (D.J. Felleman, D.C. Van Essen, 1991). Каждое из этих полей различается характером ретинотопической проекции, функциональными свойствами нейронов. К настоящему времени в иерархии корковых полей выделены и параллельные пути проведения специфической информации, например, о статических и динамических параметрах стимулов. У обезьяны такие пути (мелкоклеточные-парво-Р и крупноклеточные-магно-М) исходят от разных типов клеток сетчатки и через разные слои наружного коленчатого тела, не перемешиваясь, приходят в разные участки (модули) полей V1 и V2 (D.H. Hubel, M.S. Livingstone, 1987). Пространственное расположение этих модулей в корковых полях определено методом гистохимического выявления фермента цитохромоксидазы. Далее М и Р пути через специализированные зоны V3, V4, V5 (MT) поступают, в основном независимо, в височную и теменную кору (A. Morel, J. Bullier, 1990) . Имеются данные о конвергенции этих параллельных путей в латеральном фронтальном глазном поле коры (J. Bullier et al., 1996). К сожалению, пока не уделено должного внимания особенностям организации этих путей в разных полушариях.
Для создания образа зрительная система использует различные стратегии, комбинируя информацию из параллельных и последовательных путей (S. Zeki, S. Shipp, 1988; W.H. Merigan, J.H.R. Maunsell , 1991; D.J. Felleman, D.C. Van Essen et al., 1992; J. Bullier, 2001). Система согласованной работы двух полушарий в процессе мышления, основанная на результатах нейрофизиологических и психофизических исследований с привлечением данных клинических наблюдений, предложена В.Д. Глезером (В.Д. Глезер, 1985, V.D. Glezer, 1995).
Несомненно, функциональная межполушарная асимметрия должна быть отражена в конструкции нейронных связей корковых полей. Наиболее важными представляются связи, обеспечивающие противонаправленные потоки информации : 1) из внешнего мира, 2) из внутреннего мира, памяти. Рассмотрим основные характеристики выделяемых в коре типов нейронных связей.
В коре различают внутренние связи и внешние (P.-A. Salin, J. Bullier, 1995). Внешние связи между полями одного полушария или разных полушарий проходят в белом веществе, а, в случае межполушарных связей, и в мозолистом теле. Внутренними принято считать связи между клетками одного и того же коркового поля, образованные аксонами, не выходящими в белое вещество. Большинство внешних связей между корковыми полями двусторонние. У макак 20% связей однонаправленны, причем большинство из них нисходящие (feedback), т.е. направлены от высших структур иерархии к нижним (D.J. Felleman, D.C. Van Essen, 1991). Дисбаланс в пользу нереципрокных нисходящих связей выявлен в затылочно-височном пути (P.-A. Salin, J. Bullier, 1995).
Восходящие (feedforward) связи, по которым передается сенсорная информация, организованы ретинотопически (P.-A. Salin et al., 1989). Это означает, что рецептивное поле нейрона совпадает по размеру с совокупным рецептивным полем клеток нижележащей зрительной структуры, посылающих аксоны этому нейрону. В нисходящих корковых связях отсутствует ретинотопическое соответствие: совокупное рецептивное поле клеток вышележащего уровня превышает размер рецептивного поля клетки, в которую они проецируются (P.-A. Salin et al., 1992). Такое превышение обуславливает влияния из участков, находящихся за пределами «классического» рецептивного поля (L. Maffei, A. Fiorentini, 1976). Отличие восходящих и нисходящих связей наблюдается и в характере распределения аксонных коллатералей. В случае восходящих связей окончания коллатералей распределены в коре неравномерно, сгруппированы (patches) и контактируют с нейронами, имеющими сходные функциональные свойства. Для нисходящих связей характерно более диффузное распределение окончаний аксонных ветвей, обеспечивающее воздействия на функционально различные группы клеток. Кроме того, восходящие связи оканчиваются, в основном, в слое 4 коры, а нисходящие - во всех слоях коры, за исключением слоя 4 (D.J. Felleman, D.C. Van Essen, 1991; P.-A. Salin, J. Bullier, 1995).
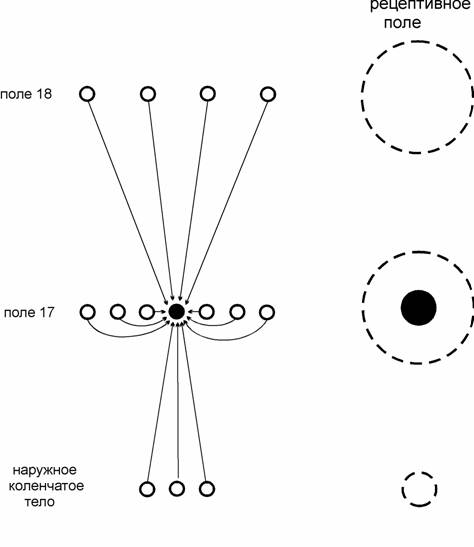
Внутренние связи полей, подобно нисходящим, организованы неретинотопически. Вследствие пачечного характера ветвлений аксонных коллатералей, они образуют системы объединений клеток со сходными функциональными свойствами, рецептивные поля которых не перекрываются (C.D. Gilbert, T.N. Wiesel, 1979, 1983; R. Malach et al., 1993). Таким образом, при посредничестве внутренних связей может происходить регулирование восходящих потоков информации нисходящими, диффузно распределенными. Пространственные соотношения восходящих, нисходящих и внутренних связей приведены схематически на рис.1. на примере коркового нейрона. Особенности организации вышеописанных типов нейронных связей в коре левого и правого полушарий до сих пор не изучены.
Рис.1. Схема конвергенции связей нейрона поля 17 коры и совокупные рецептивные поля (пунктирные окружности) конвергирующих клеток. Черные круги –нейрон поля 17 (слева) и его рецептивное поле (справа).
Пространственная организация и соотношение разных типов внутри- и межполушарных связей для полей разных иерархических уровней должны различаться исходя из следующих соображений. В разных полушариях топически организованных проекционных полей коры представлены разные половины сенсорного пространства. Биологическая значимость объектов в левой и в правой половинах пространства одинаковы, поэтому оснований для межполушарных различий в структуре и функции этих полей не имеется, структура внутренних и межполушарных связей должна быть симметрична. Наблюдаемая в первичных сенсорных полях ФМА (В.Л. Бианки, 1985), надо полагать, обусловлена несимметричной организацией нисходящих связей, обеспечивающих влияния полей более высоких уровней. По мере повышения иерархического уровня коркового поля увеличивается размер и усложняется внутренняя структура рецептивных полей нейронов (в зрительных полях эти изменения описаны как трансформации первого, второго порядка (J.M. Allman, J.H. Kaas, 1974)). Функциональные операции, осуществляемые в этих корковых полях, все более отвлечены от топографического отображения внешних воздействий, свойственного проекционным полям. Это сопровождается постепенной полушарной латерализацией этих корковых полей, которая на наивысшем уровне сводится к полностью однополушарной локализации поля, как в случае речевой зоны Брока. В связи с этим, с повышением иерархического уровня коркового поля симметричность относительно средней линии мозга организации его внутренних связей, а также межполушарных связей поля должна уменьшаться. При этом должна различаться и конструкция восходящих и нисходящих межполушарных связей этих полей в зависимости от их функциональной специализации, степени выраженности функциональной асимметрии и т.д.
В последние годы при исследовании зрительной коры крысы (J.W. Lewis, J.F. Olavarria, 1995) и кошки (J.F. Olavarria, 1996) были получены данные, противоречащие имеющимся представлениям о гомотопичности межполушарных связей в проекционных полях. Долгое время считалось, что гомотопический принцип является основным в морфологической организации всей каллозальной системы (F. Bremer, 1955; F. Ebner, R.E. Myers, 1962; G. Berlucchi, 1972). То есть, каллозальные связи связывают симметричные участки коры больших полушарий. В дальнейшем была показана неоднородность структурной организации межполушарных связей, выявлены также гетеротопические проекции между полями разных уровней и сенсорных модальностей (Ф.Н. Макаров, 1967, 1968; M.A. Segraves, A.S. Rosenquist, 1982; D. Sanides, 1978; G.M. Innocenti, 1986).
Рассмотрим более детально результаты исследований межполушарных связей зрительных проекционных полей. Из ранних морфологических исследований было известно (M.E. Wilson, 1968; G.M. Innocenti, 1986), что каллозальные и каллозально реципиентные клетки расположены вблизи границы поля 17 с полем 18. Нейрофизиологические исследования показали, что в этих участках полей представлена проекция вертикального меридиана поля зрения (D.H. Hubel, T.N. Wiesel, 1962; J. Leicester, 1968; C.J. Shatz, 1977). На основании этих данных считалось, что в мозолистом теле проходят волокна, связывающие анатомически зеркально симметричные участки полей, в которые проецируется вертикальный меридиан и близлежащая область – так называемое “правило вертикального меридиана”. Были выдвинуты 2 гипотезы о функциональной роли гомотопических каллозальных связей: 1) объединение двух половин поля зрения, 2) формирование бинокулярных рецептивных полей для стереоскопического зрения в области средней линии (D.H. Hubel, T.N. Wiesel, 1967; G. Berlucchi, G. Rizzolatti, 1968; C. Blakemore et al., 1983).
В дальнейших исследованиях на кошке было выявлено, что на границе полей 17 и 18 представлена часть ипсилатерального полуполя зрения, и эта зона названа переходной. Ширина переходной зоны 17/18 составляет около 1 мм, что соответствует 40 на уровне проекции нулевого горизонтального меридиана и около 200 на уровнях выше и ниже 200 (B.R. Payne, 1990, 1994). Следовательно, у кошки центральная часть поля зрения шириной не менее 80 представлена в коре дважды, в двух полушариях. Источником двойного представительства этой центральной части поля зрения является зона назо-темпорального перекрытия на сетчатке. Часть ганглиозных клеток из вертикальной полосы (ширина 0,2¸0,5 мм) темпоральной половины сетчатки проецируется в хиазму вместе ганглиозными клетками назальной половины (Stone, 1966), что и обеспечивает представительство ипсилатеральной половины поля зрения в переходной зоне 17/18 (B.R. Payne, 1990).
В работе Олаварриа (J.F. Olavarria, 1996) структура каллозальных связей изучалась методом локальных инъекций флуоресцентных красителей. Было показано, что при введении маркера в переходную зону 17/18 одного полушария область ретроградно меченых клеток расположена в противоположном полушарии не в переходной зоне, а вне ее. И наоборот, при введении маркера в поле 17 на расстоянии до 3 мм от переходной зоны, меченые клетки локализуются в переходной зоне 17/18 противоположного полушария. При сопоставлении местоположения каллозальных клеток с ретинотопической организацией исследованной области автор заключает, что каллозальные волокна связывают не анатомически, а ретинотопически соответствующие участки полей двух полушарий. Эти данные были подтверждены нами при исследовании каллозальных связей колонок нейронов полей 17, 18 и переходной зоны 17/18 методом микроионофоретического введения пероксидазы хрена (S.V. Alexeenko et al., 2001; С.В. Алексеенко с соавт., 2002) (рис.2).
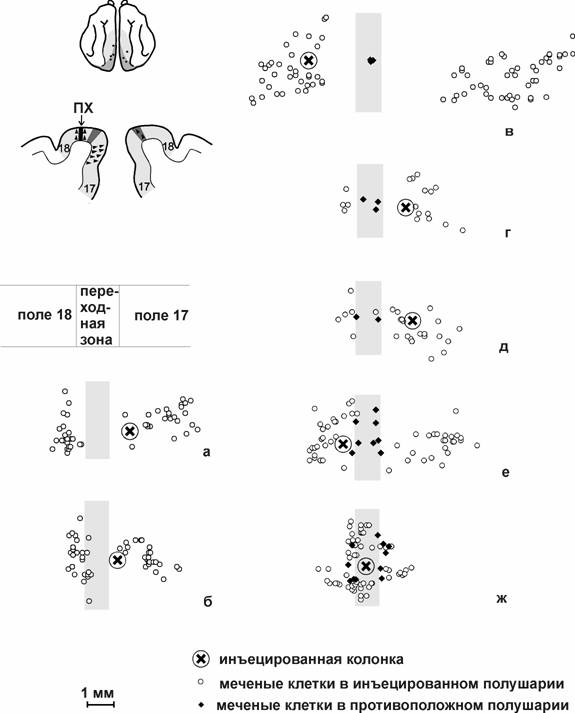
Рис.2. Распределение меченых клеток в полях 17, 18 коры кошки после введения пероксидазы хрена (ПХ) в 7 колонок нейронов. Вид сверху на поверхность коры, на которую спроецированы меченые клетки всех слоев. Распределение меченых клеток противоположного полушария наложено на распределение меченых клеток инъецированного полушария. На схеме полушарий мозга точками указана локализация колонок для а - ж. На фронтальном срезе показаны инъецированная колонка в поле 18 и меченые клетки в обоих полушариях (цит. по (Алексеенко с соавт., 2002)).
Кроме того, были выявлены колонки, не получающие каллозальных входов. По соотношению количества меченых клеток в глазоспецифичных слоях А и А1 дорзального ядра наружного коленчатого тела было определено из какого глаза исходно управляются колонки разных типов. Оказалось, что колонки полей 17 и 18 (вне переходной зоны), не получающие каллозальных входов, управляются предпочтительно из контралатерального глаза, а колонки, получающие каллозальные входы, - из ипсилатерального глаза. Колонки переходной зоны, получающие каллозальные входы, управляются из контралатерального глаза. Поиску корреляции меченых каллозальных клеток с глазодоминантными колонками коры была посвящена также и следующая работа Олаварриа (J.F. Olavarria, 2001). Для этого методом внутриглазного введения транснейронально транспортируемой пероксидазы хрена, конъюгированной с зародышем пшеницы, было визуализировано на поверхности коры распределение глазодоминантных колонок. И одновременно при введении флуоресцентных красителей выявляли каллозальные нейроны. Было показано, что каллозальные нейроны полей 17 и 18 (вне переходной зоны) локализуются, в основном, в колонках доминирования ипсилатерального глаза, а каллозальные нейроны переходной зоны 17/18 - в колонках доминирования контралатерального глаза.
Результаты этих работ свидетельствуют о зеркально симметричной относительно средней линии мозга организации прямых межполушарных связей проекционных полей коры у кошки. При этом, межполушарно связанные клетки получают вход из одного и того же глаза и локализованы в анатомически несимметричных участках разных полушарий: в переходной зоне одного полушария и в полях 17, 18 (вне переходной зоны) другого полушария. Каллозальные и каллозально-реципиентные клетки переходных зон 17/18 принадлежат к перекрещивающемуся в хиазме зрительному пути, а полей 17 и 18 - к неперекрещивающемуся пути. На рис.3 представлена схема межполушарных связей поля 18, отражающая эти данные (связи поля 17 организованы аналогично и для простоты не показаны). Такая структура прямых каллозальных связей проекционных полей коры обеспечивает первый этап реконструкции зрительного пространства - ”сшивание” полуполей зрения раздельно для каждого из глаз.
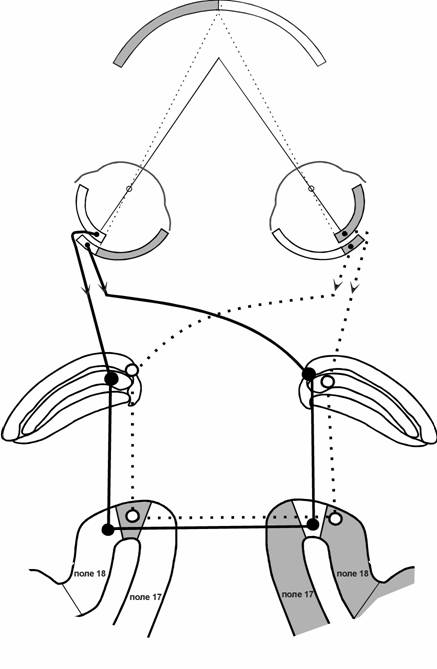
Рис.3. Схема межполушарных связей поля 18 коры кошки.
Более сложным образом должны быть организованы прямые межполушарные связи у приматов. Это обусловлено, во-первых, другой структурой зоны назо-темпорального перекрытия на сетчатке, которая является источником этих связей. У обезьян зона перекрытия на вертикальном меридиане сетчатки имеет меньшую ширину (около 10 в центре сетчатки и расширяется до 150 на крайней периферии вверху и внизу) и центрирована относительно фовеа, а не смещена в темпоральную половину, как у кошки (J. Stone, 1966; R.B. Illing, H. Wassle, 1981). Ипсилатерально проецирующиеся ганглиозные клетки окружают назальную границу фовеа, а контралатерально проецирующиеся - темпоральную границу (J. Stone et al., 1973; A.H. Bunt et al., 1977; A.G. Leventhal et al., 1988). Во-вторых, имеются сведения о том, что перекрещивающиеся в хиазме волокна из темпоральной половины сетчатки принадлежат крупноклеточному (магно) пути, а неперекрещивающиеся волокна из назальной половины – мелкоклеточному (парво) пути (Y. Fukuda et al., 1989). В третьих, в поле V2 у обезьян межполушарные связи коррелируют с распределением цитохромоксидазных полос (G.F. Olavarria, P.L. Abel, 1996), что свидетельствует о том, что распределение каллозальных связей зависит не только от топографии корковых полей, но и от распределения в них функциональных путей.
Надо полагать, что детальные исследования нейронных связей внутри и между корковыми полями разных уровней позволят определить степень их структурной симметрии.
Список литературы
1. Адрианов О.С. О принципах структурно-функциональной организации мозга // Избранные научные труды. - М. - 1999. - с. 88-96.
2. Алексеенко С.В., Топорова С.Н., Макаров Ф.Н. Нейрональные связи, обеспечивающие объединение полуполей зрения // Сенсорные системы. - 2002. - т. 16, № 2. - с.83-88.
3. Бианки В.Л. Асимметрия мозга животных. - Л: Наука. - 1985. - 295с.
4. Бианки В.Л. Механизмы парного мозга. - Л: Наука. - 1989. - 263с.
5. Боголепова И.Н. Предпосылки структурной асимметрии речедвигательной зоны Брока мозга человека // Вестник Росс. Акад.Мед.наук. – 2001. - № 4. - с. 61-64.
6. Боголепова И.Н. Показатели структурной организации некоторых корковых формаций в левом и правом полушариях мозга человека // Журн.невропатологии и психиатрии им.Корсакова. – 1981. - т. 81, № 7. - с. 974-977.
7. Болотов А.В. Функциональная асимметрия 21 поля у кошки // Матер. конф. ”Актуальные вопросы функциональной межполушарной асимметрии”. - М. – 2001. - с. 26-27.
8. Габибов И.М., Глезер В.Д. Асимметрия размеров рецептивных полей нейронов 21-го поля коры больших полушарий мозга кошки // Докл.АН СССР. – 1985. - т. 284, № 4. – с. 1014-1015.
9. Глезер В.Д Зрение и мышление. - Л: Наука. – 1985. –246 с.
10. Кононова Е.П. Вариабельность строения коры большого мозга. Нижняя фронтальная извилина у человека // В кн.: Труды Ин-та мозга.- М.-Л. – 1935. - в.1. - с. 50-118.
11. Леушина Л.И., Невская А.А., Павловская М.Б. Асимметрия полушарий головного мозга с точки зрения опознания зрительных образов // В кн.: Сенсорные системы. Зрение. – Л. – 1982. - с. 76-92.
12. Макаров Ф.Н. Гетеротопические комиссуральные связи 17-го поля кошки // ДАН СССР. – 1967. - т. 176, № 5. - с. 1192-1194.
13. Макаров Ф.Н. Комиссуральные связи зрительной коры кошки (17, 18, 19 поля) // Арх.анат., гист. и эмбриол. – 1968. - т. 55, вып. 7. - с. 48-54.
14. Меерсон Я.А. Высшие зрительные функции. - Л. – 1989. – 163 с.
15. Невская А.А., Леушина Л.И. Асимметрия полушарий и опознание зрительных образов. – Л. – 1990. – 152с.
16. Праздникова Н.В., Глезер В.Д., Макаров Ф.Н. О механизмах инвариантного описания образа и полного описания изображения в левом и правом полушариях кошки // Сенс. Сист. – 1997. – т.11, № 3. - с.352–359.
17. Праздникова Н.В., Глезер В.Д., Гаузельман В.Е. О механизмах определения порогов в левом и правом полушариях кошки // Сенс. Сист. – 1999. – т.13, № 2. - с.152–157.
18. Супин А.Я. Нейрофизиология зрения млекопитающих. - М.: Наука. – 1981. - 252 с.
19. Alexeenko S.V., Toporova S.N., Makarov F.N. The fine structure of interhemispheric connections in areas 17,18 of the cat // Perception. – 2001. - vol. 30. - p. 115.
20. Allman J.M., Kaas J.H. The organization of the second visual area (VII) in the owl monkey: the second order transformation of the visual hemifield // Brain Res. - 1974, - vol .76. - p. 247-265.
21. Blakemore C., Diao Y., Pu M., Wang Y., Xiao Y. Possible functions of the interhemispheric connections between visual cortical areas in the cat // J.Physiol.(London). – 1983. - vol. 337. - p. 334-349.
22. Berlucchi G. Anatomical and physiological aspects of visual functions in the corpus callosum // Brain Res. – 1972. - vol. 37. - p. 371-392.
23. Berlucchi G., Rizzolatti G. Binocularly driven neurons in visual cortex of split chiasm cats // Science. – 1968. - vol. 159. - p. 308-310.
24. Bremer F. Responses of visual area to callosal impulses in the cat // Proc.Soc.Exp. Biol. Med. – 1955. - vol. 90. - p. 22-25.
25. Bullier J. Integrated model of visual processing // Brain Res Rev. – 2001. – vol. 36, № 2-3. – p. 96-107.
26. Bullier J, Schall JD, Morel A. Functional streams in occipito-frontal connections in the monkey // Behav Brain Res. – 1996. – vol. 76, № 1-2. – p. 89-97.
27. Bunt A. H., Minckler S., Johanson G.W. Demonstration of bilateral projection of the central retina of the monkey with horseradish peroxidase neuronography // J.Comp.Neurol. - 1977. – vol.171, №4. - p. 619-630.
28. Christman S. Perceptual characteristics in visual laterality research // Brain and cognition. – 1989. - vol. 11. - p. 238-257.
29. Ebner F., Myers R.E. Corpus callosum and interhemispheric transmission of actual learning // J.Neurophysiol. – 1962. - vol. 25. - p. 380-391.
30. Felleman D.J., Van Essen D.C. Distributed hierarchical processing in the primate cerebral cortex // Cerebral Cortex. - 1991. - № 1. - p. 1-47.
31. Fukuda Y., Sawai H., Watanabe M., Morigiwa K. Nasotemporal overlap of crossed and uncrossed retinal ganglion cell projections in the Japanese monkey (Macaca fuscata) // J.Neurosci. – 1989. – vol. 9, № 7. – p.2353-2373
32. Galaburda A.M., LeMay M., Kemper T.L., Geschwind N. Right-left asymmetries in the brain // Science. – 1978. - vol.199. - p. 852-856.
33. Geschwind N., Levitsky W. Human brain: left-right asymmetries in temporal speech region // Science. – 1968. - vol.161. - p. 186-187.
34. Gilbert C.D., Wiesel T.N. Morphology and intracortical projections of functionally characterized neurons in the cat visual cortex // Nature.- 1979. - vol. 280.- p. 120-125.
35. Gilbert C.D, Wiesel T.N. Clustered intrinsic connections in cat visual cortex // J.Neurosci. - 1983. - vol. 3, № 4.- p. 1116-1133.
36. Glezer V.D. Vision and mind:modelling mental functions. – Lawrens Erlbaum Ass.Publ., Mahwah N.J. –1995. –274p.
37. Hubel D.H., Wiesel T.N. Cortical and callosal connections concerned with the vertical meridian of visual fields in the cat // J.Neurophysiol. – 1967. - vol.30, № 6. - p. 1561-1573.
38. Hubel D.H., Wiesel T.N. Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex // J.Physiol.(Lond). – 1962. - vol. 160. – p. 106-154.
39. Hubel D.H., Livingstone M.S. Segregation of form, color, and stereopsis in primate area 18 // J. Neurosci. - 1987. - vol. 7, № 11. - p. 3378-3415.
40. Innocenti G.M. General organization of callosal connections in the cerebral cortex // In: Cerebral Cortex, ed. E.G.Jones, A.Peters. - New York:Plenum. – 1986. - vol. 5. - p. 291-351.
41. Illing R.B., Wassle H. The retinal projection to the thalamus in the cat: a quantitative investigation and a comparison with the retinotectal pathway // J.Comp.Neurol. - 1981. - vol. 202. – p. 265-285.
42. Leicester J. Projection of the vertical meridian to cerebral cortex of the cat // J.Neurophysiol. 1968. - vol.31, № 3. - p. 371-382.
43. Leventhal A.G., Ault S.J., Vitek D.J. The nasotemporal division in primate retina: the neural bases of macular sparing and splitting // Science. – 1988. – vol. 240, № 4848. – p. 66-67.
44. Lewis J.W., Olavarria J.F. Two rules for callosal connectivity in striate cortex of the rat // J.Comp.Neurol. – 1995. - vol. 361, №1. - p.119-137.
45. Maffei L., Fiorentini A. The unresponsive regions of visual cortical receptive fields // Vision Res. – 1976. - vol. 16, - p. 1131-1139.
46. Malach R., Amir Y., Harel M., Grinwald A. Relationship between intrinsic connections and functional architecture revealed by optical imaging and in vivo targeted biocytin injections in primate striate cortex // Proc.Natl.Acad.Sci.USA.- 1993.- vol. 22. - p. 10469-10473.
47. Merigan W.H., Maunsell J.H.R. How parallel are the primate visual pathways? // Ann.Rev.Neurosci. – 1991. - vol. 16. - p. 369-402.
48. Morel A, Bullier J. Anatomical segregation of two cortical visual pathways in the macaque monkey // Vis Neurosci. – 1990. - vol.4, №6. –p. 555-578.
49. Olavarria J.F. Non-mirror-symmetric patterns of callosal linkages in areas 17 and 18 in cat visual cortex // J. Comp. Neurol. - 1996. - vol. 366. -p. 643-655.
50. Olavarria J.F. Сallosal connections correlate preferentially with ipsilateral cortical domains in cat areas 17 and 18, and with contralateral domains in the 17/18 transition zone // J. Comp. Neurol. - 2001. - vol. 433. - p.437-457.
51. Olavarria J.F., Abel P.L. The distribution of callosal connections correlates with the pattern of CO stripes in visual area V2 of macaque monkey // Cerebral Cortex. – 1996. - vol. 6, №4. - p. 631-639.
52. Orban G.A. Neuronal operations in the visual cortex. - Berlin. Springer-Verlag. - 1984. – 367p.
53. Payne B.R. Representation of the ipsilateral visual field in the transition zone between areas 17 and 18 of the cat’s cerebral cortex // Visual Neurosci. – 1990. - vol. 4, № 3. - p. 445-474.
54. Payne B.R. Neuronal interactions in cat visual cortex mediated by the corpus callosum // Behav. Brain Res. – 1994. - vol. 64. - p.55-64.
55. Rosen G.D. Cellular, morphometric, ontogenetic and connectional substrates of anatomical asymmetry // Neurosci.and Biobehav. Rev. – 1996. - vol.20, № 4. - p. 607-615.
56. Salin P.-A., Bullier J. Corticocortical connections in the visual system: structure and function // Physiol. Rev. - 1995. - vol. 75, №1. - p. 107-154.
57. Salin P.-A., Bullier J., Kennedy H. Convergence and divergence in the afferent projections to cat area 17 // J. Comp. Neurol. - 1989.- vol. 283, №4. – p. 486-512.
58. Salin P.-A., Girard P., Kennedy H., Bullier J. Visuotopic organization of corticocortical connections in the visual system of the cat // J. Comp. Neurol. - 1992. - vol. 320, №4. - p. 415-454.
59. Sanides D. The retinotopic distribution of visual callosal projections in suprasylvian visual area compared to the classical visual areas (17, 18, 19) in the cat // Exp.Brain Res. – 1978. - vol.33, - p. 435-443.
60. Scannell J.W., Blakemore C., Young M.P. Analysis of connectivity in the cat cerebral cortex // J.Neurosci. - 1995. - vol. 15, № 2. - p. 1463-1483.
61. Segraves M.A., Rosenquist A.C. The distribution of the cells of origin of callosal projections in cat visual cortex // J. Neurosci.. – 1982. - vol.2. – p.1079-1089.
62. Shatz C. J. Anatomy of interhemispheric connections in the visual system of Boston Siamese and ordinary cats // J.Comp. Neurol. – 1977. - vol. 173. - p. 497-518.
63. Stone J. The naso-temporal division of the cat’s retina // J.Comp.Neurol. – 1966. - vol.126, N4. – p. 585-599.
64. Stone J., Leicester J., Sherman S.M. The naso-temporal division of the monkey’s retina // J.Comp.Neurol.- 1973. - vol.150, №3. - p.333-348.
65. Van Essen D.C., Anderson C.H., Felleman D.J. Information processing in the primate visual system: an integrated systems perspective // Science. – 1992. - vol. 255, №5043. - p. 419-423.
66. Wilson M.E. Cortico-cortical connexions of the cat visual areas // J.Anat. – 1968. – vol. 102. – p. 375-386.
67. Zeki S., Shipp S. The functional logic of cortical connections // Nature. - 1988. - vol. 335. – p. 311-317.